

Запас питательных веществ: Запасание питательных веществ — урок. Биология, Бактерии. Грибы. Растения (5–6 класс).
Запасание питательных веществ — урок. Биология, Бактерии. Грибы. Растения (5–6 класс).
Часть органических веществ не сразу расходуется на процессы жизнедеятельности, а сохраняется растением про запас.
У однолетних растений питательные вещества запасаются в плодах и семенах.
Рис. \(1\). Плоды тыквы
У двулетних и многолетних растений местами хранения запасов органических веществ служат стебли, корни и их видоизменения. Корнеплоды, клубни и луковицы — это хранилища запасных органических веществ.
Рис. \(2\). Корнеплоды
Пример запасания веществ в стебле — утолщённый разросшийся стебель капусты кольраби.
Рис. \(3\). Кольраби
Кустарники и деревья запасают питательные вещества в древесине и сердцевине.
Рис. \(4\). Слои ствола
С наступлением весны запасные вещества переходят в растворимое состояние и вместе с водой поступают к распускающимся почкам. Многие пили берёзовый или кленовый сок. Сок собирают ранней весной. Аккуратно делают отверстие в коре дерева и капли сока медленно стекают в приготовленный сосуд.
Рис. \(5\). Сбор берёзового сока
Собирать сок нужно аккуратно, стараясь не причинить вред дереву. Если дерево потеряет много сока, то оно будет плохо расти и может погибнуть.
Источники:
Рис. 1. Плоды тыквы. https://cdn.pixabay.com/photo/2017/11/30/22/00/pumpkin-2989569_960_720. 21.10.2021.
Рис. 2. Корнеплоды. https://cdn.pixabay.com/photo/2016/09/01/03/25/vegetables-1635167_960_720. 21.10.2021.
Рис. 3. Кольраби. https://image.shutterstock.com/image-photo/gardening-fresh-kohlrabi-turnip-600w-107637092
Рис. 4. Слои ствола. https://image.shutterstock.com/image-vector/illustration-biology-anatomy-trunk-tree-600w-1441257410
Рис. 5. Сбор берёзового сока. https://image.shutterstock.com/image-photo/collecting-birch-sap-wooden-pipe-600w-399784282
5. Сбор берёзового сока. https://image.shutterstock.com/image-photo/collecting-birch-sap-wooden-pipe-600w-399784282
Животные потеряли способность производить желток около 30–70 миллионов лет назад
Животные долго не доверяли молоку заботу о потомстве. Как показали швейцарские учёные, 200 миллионов лет они сохраняли запасной вариант — накопление желтка в яйцеклетке, и лишь 30–70 миллионов лет назад наши предки окончательно лишились такой возможности.
Омлет — очень питательный завтрак, но большинство всё же предпочитают либо молоко, либо яйца. Такой же выбор сделала природа, лишившая млекопитающих возможности вскармливать детей желтками около 30–70 миллионов лет назад. Швейцарские учёные смогли шаг за шагом проследить, как животные перешли к кормлению детёнышей исключительно молоком — стратегии, используемой самыми успешными на сегодняшний день представителями фауны.
На эмбриональной стадии развивающемуся организму нужно особенно много питательных веществ и энергии, ведь за считанные дни одна клетка увеличивается в десятки и сотни раз. За долгие миллионы лет эволюции природа испробовала разные варианты обеспечения этой потребности.
За долгие миллионы лет эволюции природа испробовала разные варианты обеспечения этой потребности.
Самые древние организмы ограничивались тем, что оставляли детёнышам запас питательных веществ раз и навсегда, накапливая в яйцеклетке достаточно «пищи» для формирования полноценного организма. Если у яйцеклетки есть твёрдая оболочка, то мы обычно называем её яйцом, а запас питательных веществ в ней — яичным желтком.
Яйцеклетки и желток
Яйцеклетки — гаплоидные клетки, являющиеся женскими гаметами у человека, животных и семянных растений, образуются в результате овогенеза. Человеческая яйцеклетка имеет диаметр примерно 145 микрон и является самой большой клеткой…
Чем большего размера появляющийся на свет детёныш, тем больше веществ приходится откладывать. Например, ооцит — материнская половая клетка — некоторых акул достигает 22 см в поперечнике. Однако такая тактика позволяет, однажды накопив питательные вещества, больше не заботиться о развитии зародыша — на свет появляется организм, способный питаться уже, как взрослый, пусть и не без помощи родителей, ковыряющих ему червячков.
Другая крайность — непрерывно заботиться о зародышах с самого начала, начиная практически с момента зарождения жизни — формирования зиготы. После появления детёныша на свет такие животные продолжают кормить его — теперь уже материнским молоком. В числе этих самых совершенных животных и человек.
Как показали швейцарцы, переход от одной крайности к другой продолжался более 200 миллионов лет, а материнское молоко появилось задолго до того, как яйцеклетки лишились своих желтков.
Работа Давида Браванда, Вальтера Вали и Хенрика Каэссманна из Университета Лозанны опубликована в последнем номере PLoS Biology.
Учёные использовали тот факт, что среди млекопитающих есть три группы, использующие разную стратегию выведения потомства. Это плацентарные, к которым относится большинство, в том числе человек, кошка, собака; сумчатые, распространенные в Австралии и островах Океании, а также в Южной Америке; и однопроходные — утконос и пара видов ехидн.
Самки плацентарных длительное время вынашивают эмбрион внутри, за чем следует относительно короткий период лактации, не превышающий время эмбрионального развития.
У сумчатых плацента развита слабо и не может прокормить большой эмбрион, а потому период беременности небольшой, и за ним следует продолжительное вскармливание. Например, у кенгуру, которые могут быть значительно больше человека, беременность длится всего несколько недель, а потом очень маленький малыш, вес которого не превышает 1 грамма, переползает в сумку, где и питается молоком до 1 года. Все, чем мать может помочь ему — вылизать дорогу, облегчив путь по шерсти.
Первозвери (Prototheria)
подкласс примитивных млекопитающих, соединяющих в себе черты млекопитающих, рептилий и птиц. В этом подклассе выделяют единственный инфракласс клоачные, противопоставляемый инфраклассам плацентарные и сумчатые из подкласса звери…
Однопроходные проводят с потомством еще меньше времени. Как последние, сохранившиеся из первозверей, они откладывают яйца, похожие на рептилий. У утконоса уже через 10 дней вылупляются детеныши размером до 2,5 см, которые потом питаются молоком матери, выступающим из пор на животе и стекающим по складкам. И уже в 17 недель они покидают нору и идут на свою первую охоту.
И уже в 17 недель они покидают нору и идут на свою первую охоту.
Всё это многообразие произошло от рептилий, зачастую даже не заботящихся о высиживании яиц. Ученые сравнили гены представителей трех этих групп животных с «контролем» — курицей, как и все птицы, тоже произошедшей от яйцекладущих чешуйчатых предков и к сегодняшнему дню ни утерявшей способности нести яйца, ни научившейся вскармливать птенцов молоком.
close
100%
Общие участки ДНК у всех групп млекопитающих позволили ученым сказать, что гены казеина — основного белка молока — возникли между 200 и 310 миллионами лет назад, что соответствует палеонтологическим данным.
Однако палеонтология до сих пор не могла дать ответа на вопрос, когда яйцеклетки млекопитающих лишились своих желтков. Через миллионы лет до нас доходят только кости, да и то — лишь их форма — оригинальный костный материал при окремнении замещается камнем.
Чтобы выяснить, когда у яиц не стало желтков, учёные провели анализ генов, отвечающих за производство белка вителлогенина — основного источника питания в яйце.
У курицы таких генов три. У однопроходных — самых близких к древнейшим предкам животных — ученые нашли только один. Остальные два превратились в псевдогены — участки ДНК, потенциально несущие ту же информацию и даже представленные почти той же последовательностью, за исключением нескольких мутаций, которые и сделали их неактивными. Переход от гена к псевдогенам проходил постепенно, и последняя мутация, сделавшая ген окончательно бесполезным, случилась 30–70 миллионов лет назад.
В результате, если утконос ещё может обеспечить своих потомков небольшим запасом желтка, то размер человеческой яйцеклетки (кстати, самой большой клетки в человеческом организме) не превышает 145 мкм.
Обнаружить животных, у которых активны два гена виттелогенина, пока не удалось. Не исключено, что они просто не сохранились к настоящему времени — ведь и тот факт, что на нашей планете до сих пор живут утконос и ехидна, многие учёные считают чудом.
Всё, что ни происходит, всё к лучшему. И если эволюция и лишила нас желтка, то дала потрясающий источник энергии и питательных веществ, а это позволяет сочетать знакомство с большим миром с ростом и развитием, не ограниченным скорлупой.
Запас питательных веществ — это… Что такое Запас питательных веществ?
- Запас питательных веществ
- — валовое содержание питательных веществ в определенном слое п. Выражается в кг/га.
Толковый словарь по почвоведению. — М.: Наука.
Под редакцией А.А. Роде.
1975.
- Запас влаги мертвый
- Запас солей
Смотреть что такое «Запас питательных веществ» в других словарях:
-
Передвижение веществ в растении — а) П. сырого сока. Для растения необходима вода и некоторые минеральные вещества. Первая нужна как для питания, так и потому, что растение широко пользуется ею в качестве строительного материала: дряблый вид завядшего, т. е. потерявшего часть… … Энциклопедический словарь Ф.А. Брокгауза и И.
 А. Ефрона
А. Ефрона -
ЦВЕТКОВЫЕ — покрытосеменные (Angiospermae), самый крупный отдел царства растений, характеризующийся специализированными органами размножения, образующими цветок. Цветковые растения известны с юрского периода (примерно 150 млн. лет назад): уже в то время они… … Энциклопедия Кольера
-
СЕМЯ — зародышевая стадия семенного растения, образующаяся в процессе полового размножения и служащая для расселения. Внутри семени находится зародыш, состоящий из зародышевых корешка, стебелька и одного или двух листьев, или семядолей. Цветковые… … Энциклопедия Кольера
-
Подотряд Разноядные жуки (Polyphaga) — Этот подотряд гораздо обширнее первого. Как это отражено в названии подотряда, пищевые связи его представителей могут быть самыми разнообразными. Он включает основную массу жесткокрылых и делится на большое число семейств.… … Биологическая энциклопедия
-
Ожирение — Степени ожирения: силуэты людей с нормальной и избыточной … Википедия
-
Насекомые* — (In secta s.
 Hexapoda) составляют один из классов типа суставчатоногих (членистоногих; Arthropoda), подтипа трахейных (Tracheata). Они могут быть коротко охарактеризованы, как трахейные суставчатоногие с телом, разделенным на голову, грудь и… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Hexapoda) составляют один из классов типа суставчатоногих (членистоногих; Arthropoda), подтипа трахейных (Tracheata). Они могут быть коротко охарактеризованы, как трахейные суставчатоногие с телом, разделенным на голову, грудь и… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона -
Насекомые — (In secta s. Hexapoda) составляют один из классов типа суставчатоногих (членистоногих; Arthropoda), подтипа трахейных (Tracheata). Они могут быть коротко охарактеризованы, как трахейные суставчатоногие с телом, разделенным на голову, грудь и… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
-
Корень — У этого термина существуют и другие значения, см. Корень (значения) … Википедия
-
Корень (биология) — Корень осевой, подземный вегетативный орган высших растений, обладающий неограниченным ростом в длину и положительным геотропизмом. Корень осуществляет закрепление растения в почве и обеспечивает поглощение и проведение воды с растворёнными… … Википедия
-
Корень (ботаника) — Корень осевой, подземный вегетативный орган высших растений, обладающий неограниченным ростом в длину и положительным геотропизмом.
 Корень осуществляет закрепление растения в почве и обеспечивает поглощение и проведение воды с растворёнными… … Википедия
Корень осуществляет закрепление растения в почве и обеспечивает поглощение и проведение воды с растворёнными… … Википедия
Как фермеры разных стран выращивают продовольственные культуры? — Конвенция о биологическом разнообразии
Как фермеры разных стран выращивают продовольственные культуры?
Выращивать зерновые без опылителей трудно, если вообще возможно, даже при наличии природных богатств, энергии, инструментов и денег. Опылители оплодотворяют цветущие зерновые, перенося пыльцу с одного цветка на другой.Опылять цветы могут насекомые, птицы, мелкие млекопитающие и даже ветер. Чаще всего опылителем являются пчелы; благодаря пчелам из цветков кофе вырастают кофейные бобы в Колумбии, из цветков манго — плоды манго в Индии, из цветков маракуйи — плоды маракуйи в Бразилии и из цветков арбуза — плоды арбузов в Китае. И не забывайте, что пчелы еще дают мед.
Задавались ли вы когда-либо вопросом о том, как фермерам удается выращивать здоровые зерновые на одном участке земли год за годом, сохраняя при этом в почве запас питательных веществ? Круговорот питательных веществ — важная разновидность эксплуатации экосистемы, благодаря которой питательные вещества используются несколькими организмами и позже возвращаются в почву, чтобы начать новый цикл. В Южной Африке фермеры для выпаса приводят своих овец на большие пастбища. Травы и цветы на пастбищах через свои корни забирают питательные вещества из почвы. Различные виды трав и растений впитывают различные питательные вещества и перерабатывают их в витамины и минералы. Когда овца ест растения, питательные вещества поступают в ее организм и используются для роста. Некоторые питательные вещества не будут впитываться в организме животного и просто пройдут через пищеварительную систему. Через некоторое время черви, бактерии и микроорганизмы расщепят питательные вещества в овечьих экскрементах, чтобы превратить их в удобрения или навоз, и питательные вещества вернутся в почву. Теперь питательные вещества доступны для трав и цветов, и может начаться новый цикл.
В Южной Африке фермеры для выпаса приводят своих овец на большие пастбища. Травы и цветы на пастбищах через свои корни забирают питательные вещества из почвы. Различные виды трав и растений впитывают различные питательные вещества и перерабатывают их в витамины и минералы. Когда овца ест растения, питательные вещества поступают в ее организм и используются для роста. Некоторые питательные вещества не будут впитываться в организме животного и просто пройдут через пищеварительную систему. Через некоторое время черви, бактерии и микроорганизмы расщепят питательные вещества в овечьих экскрементах, чтобы превратить их в удобрения или навоз, и питательные вещества вернутся в почву. Теперь питательные вещества доступны для трав и цветов, и может начаться новый цикл.
Знаете ли вы,
— что рою пчел нужно пролететь 55 тыс. миль, чтобы собрать 1 фунт меда?
— что пчела может летать со скоростью 15 миль/час?
— что в мире существует 914 пород овец и что более 85 из них выращивают в Великобритании?
§35.
 Половое размножение. Образование половых клеток
Половое размножение. Образование половых клеток
1. Как называются органы, в которых осуществляется образование женских и мужских гамет у споровых растений? У животных?
Яичники, антеридии, спорангии, семенники, архегонии.
У споровых растений женские гаметы образуются в архегониях, а мужские – в антеридиях.
У животных женские гаметы формируются в яичниках, а мужские – в семенниках.
2. Как строение сперматозоида и яйцеклетки связано с функциями, выполняемыми этими клетками?
Сперматозоиды и яйцеклетки – половые клетки, посредством которых осуществляется передача наследственной информации от одного поколения другому. Выполнение этой функции обусловлено наличием ядра с гаплоидным набором хромосом как в мужских, так и в женских гаметах. При слиянии сперматозоида и яйцеклетки образуется диплоидная зигота, из которой развивается организм, унаследовавший уникальную комбинацию генов и признаков обоих родителей.
Сперматозоиды – подвижные половые клетки. В связи с этим они имеют обтекаемую форму и длинный жгутик (хвост), служащий для активного передвижения. Сперматозоид содержит небольшое количество цитоплазмы с минимальным набором органоидов. На переднем конце головки расположена акросома, обеспечивающая растворение оболочек яйцеклетки при оплодотворении. В шейке располагаются две центриоли, одна из которых служит центром организации микротрубочек, образующих подвижную осевую нить жгутика. Митохондрии, расположенные в средней части сперматозоида, генерируют энергию, необходимую для движения жгутика.
Яйцеклетки чаще всего неподвижны. Они содержат запас питательных веществ, необходимых для развития зародыша. Поэтому яйцеклетки, как правило, намного крупнее сперматозоидов и покрыты несколькими защитными оболочками.
3. Сперматозоиды практически не содержат цитоплазмы и питательных веществ, однако им необходимо большое количество энергии для движения. Как вы думаете, откуда берётся эта энергия?
Как вы думаете, откуда берётся эта энергия?
Энергию для движения жгутика сперматозоида (в виде АТФ) непосредственно поставляют митохондрии, локализованные в средней части сперматозоида. Однако исходным источником энергии, за счёт которого происходит синтез АТФ в митохондриях, служат питательные вещества (прежде всего – фруктоза), содержащиеся в семенной жидкости. Эти вещества выделяются семенными пузырьками, предстательной железой и некоторыми другими органами мужской половой системы.
4. Какое максимальное количество яйцеклеток и вторичных полярных телец может сформироваться у кошки из четырёх ооцитов первого порядка?
У млекопитающих в результате мейотического деления ооцита первого порядка образуется одна яйцеклетка и три вторичных полярных тельца. Следовательно, из четырёх ооцитов первого порядка может сформироваться четыре яйцеклетки и двенадцать вторичных полярных телец.
5. Какие процессы, происходящие в ходе оогенеза, обеспечивают накопление в яйцеклетках большого количества питательных веществ?
Какие процессы, происходящие в ходе оогенеза, обеспечивают накопление в яйцеклетках большого количества питательных веществ?
В период роста происходит значительное увеличение объёма первичных половых клеток (оогониев) за счёт поступления в них питательных веществ из окружающих фолликулярных клеток и крови. Так образуются ооциты первого порядка.
В результате последующего мейоза (период созревания) образуются разные по величине дочерние клетки: одна крупная яйцеклетка и три небольших полярных тельца, которые впоследствии погибают и разрушаются. Биологический смысл такого неравномерного деления заключается в сохранении в яйцеклетке максимального количества питательных веществ, необходимых для развития будущего зародыша.
6. Каков биологический смысл образования при оогенезе полярных телец?
Формирование полярных телец, содержащих незначительное количество цитоплазмы, необходимо для осуществления мейоза при сохранении максимального запаса питательных веществ в яйцеклетке. Проще говоря, полярные тельца образуются с целью отделения (удаления) «избыточной» генетической информации.
Проще говоря, полярные тельца образуются с целью отделения (удаления) «избыточной» генетической информации.
7. Сравните процессы сперматогенеза и оогенеза, укажите черты сходства и различия.
Сходство:
● Представляют собой процессы гаметогенеза – образования и развития половых клеток (гамет), подразделяются на несколько периодов, протекающих сходным образом.
● Первый этап – период размножения. Диплоидные предшественники гамет многократно делятся митозом, в результате чего их количество значительно возрастает.
● Второй этап – период роста. Сперматогонии (оогонии) увеличиваются в размерах и становятся сперматоцитами (ооцитами) первого порядка.
● Третий этап – период созревания. Происходит мейоз, в результате которого из каждого сперматоцита (ооцита) первого порядка, содержащего набор 2n4c, образуются четыре гаплоидных дочерних клетки (набор 1n1c).
Различия:
● В результате сперматогенеза образуются подвижные мужские половые клетки – сперматозоиды, в результате оогенеза – неподвижные женские гаметы – яйцеклетки. Обычно сперматозоидов в мужском организме формируется во много раз больше, чем яйцеклеток в женском.
Обычно сперматозоидов в мужском организме формируется во много раз больше, чем яйцеклеток в женском.
● В сперматогенезе за периодом созревания следует период формирования сперматозоидов, в оогенезе период формирования отсутствует. Таким образом, сперматогенез включает четыре периода, а оогенез – три.
● Сперматогенез у самцов млекопитающих начинается с периода полового созревания. Оогенез у самок млекопитающих начинается ещё до рождения.
● В период роста сперматогонии увеличиваются в размерах незначительно, а размер оогониев увеличивается существенно.
● В ходе сперматогенеза мейоз приводит к образованию из одного сперматоцита первого порядка четырёх одинаковых дочерних клеток – сперматид. В ходе оогенеза происходит неравномерное мейотическое деление ооцита первого порядка, в результате чего формируется одна крупная клетка с запасом питательных веществ (яйцеклетка) и три мелких полярных тельца.
…и (или) другие существенные признаки.
8. В яичниках 22-летней женщины со стабильным 28-дневным репродуктивным циклом содержится 42 тысячи фолликулов. Большинство из них очень мелкие, и лишь 299 имеют диаметр свыше 100 мкм. Кроме того, в яичниках есть 5 жёлтых тел и 112 рубцов, оставшихся от них. В каком возрасте у этой женщины произошла первая овуляция? В каком возрасте у неё, вероятнее всего, прекратится образование яйцеклеток?
● Из 42 тысяч фолликулов только 299 имеют диаметр свыше 100 мкм, т.е. содержат ооциты первого порядка. Остальные фолликулы содержат оогонии (не прошедшие период роста) и впоследствии подвергнутся дегенерации.
● При стабильном 28-дневном репродуктивном цикле в год происходит 365 : 28 = 13,035 ≈ 13 овуляций. Общее количество жёлтых тел и рубцов, оставшихся от них, равно 5 + 112 = 117. Следовательно, первая овуляция, скорее всего, произошла 117 : 13 = 9 лет тому назад. Сейчас женщине 22 года, значит, овуляции у неё начались в 22 – 9 = 13 лет.
● Запаса ооцитов первого порядка может хватить на 299 : 13 = 23 года. Таким образом, образование яйцеклеток, вероятнее всего, прекратится в 22 + 23 = 45 лет.
Ответ: первая овуляция произошла в 13 лет; образование яйцеклеток, вероятнее всего, прекратится в 45 лет.
Дашков М.Л.
Сайт: dashkov.by
Вернуться к оглавлению
| < Предыдущая | Следующая > |
|---|
Строение семян | Параграф 18
«Биология. Бактерии, грибы, растения. 6 класс». В.В. Пасечник
Вопрос 1. Какие растения называют двудольными, а какие — однодольными?
У двудольных растений зародыши семян имеют две семядоли (фасоль, горох, яблоня и многие другие). Зародыши семян однодольных растений имеют одну семядолю (пшеница, кукуруза и др.).
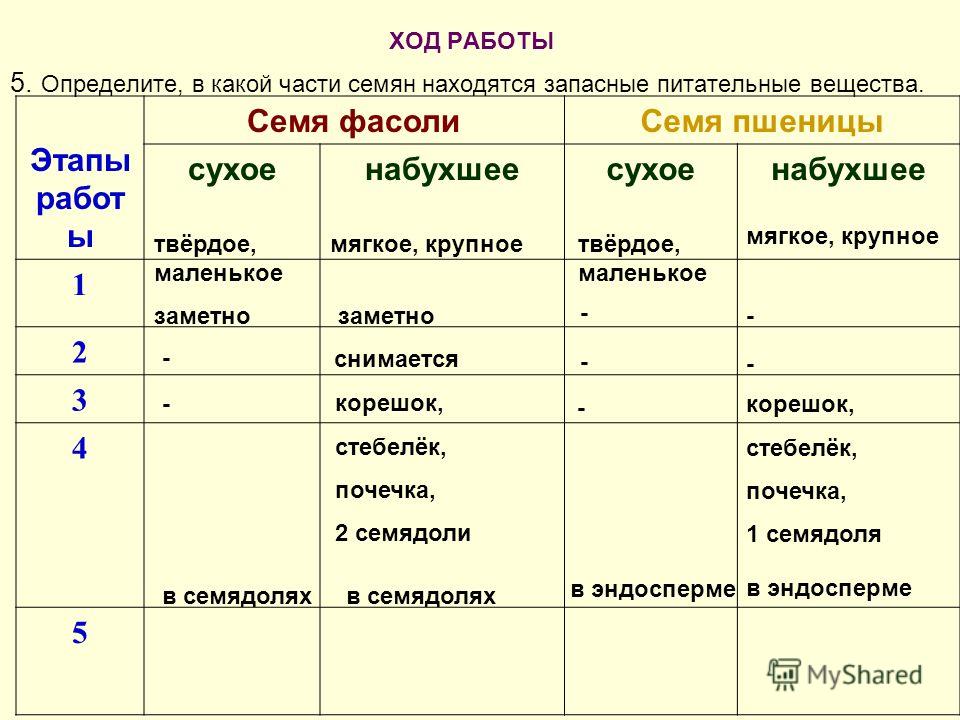
Вопрос 2. Каково строение семени фасоли?
Строение семени двудольного растения рассмотрим на примере семени фасоли (рис. 1). Снаружи семя покрыто гладкой семенной кожурой, которая образовалась из покровов семязачатка. Семенная кожура защищает семя от потерь влаги и механических повреждений. На вогнутой стороне семени заметен рубчик — след от семяножки, при помощи которой семя крепилось к околоплоднику. Под семенной кожурой располагается зародыш. Зародыш состоит из двух мясистых семядолей, в которых сосредоточен запас питательных веществ, зародышевых корешка, стебелька и почечки. На вогнутой стороне семени имеется рубчик — место прикрепления семени к семяножке. Над рубчиком находится маленькое отверстие — микропиле. Через микропиле в семя проникают воздух и вода. Семя фасоли не содержит эндосперма. Запасные питательные вещества сосредоточены в зародышевых стебельке, корешке, а также семядолях, поэтому перечисленные части зародыша выполняют питательную функцию.
1). Снаружи семя покрыто гладкой семенной кожурой, которая образовалась из покровов семязачатка. Семенная кожура защищает семя от потерь влаги и механических повреждений. На вогнутой стороне семени заметен рубчик — след от семяножки, при помощи которой семя крепилось к околоплоднику. Под семенной кожурой располагается зародыш. Зародыш состоит из двух мясистых семядолей, в которых сосредоточен запас питательных веществ, зародышевых корешка, стебелька и почечки. На вогнутой стороне семени имеется рубчик — место прикрепления семени к семяножке. Над рубчиком находится маленькое отверстие — микропиле. Через микропиле в семя проникают воздух и вода. Семя фасоли не содержит эндосперма. Запасные питательные вещества сосредоточены в зародышевых стебельке, корешке, а также семядолях, поэтому перечисленные части зародыша выполняют питательную функцию.
Рис. 1. Строение семени фасоли.
Вопрос 3. Где находится запас питательных веществ в семенах фасоли, ясеня, миндаля?
У фасоли запас питательных веществ находится в клетках зародыша, в основном в семядолях. У ясеня и миндаля запас питательных веществ находится в особой запасающей ткани — эндосперме, который располагается под семенной кожурой и окружает зародыш будущего растения.
Вопрос 4. Какое строение имеет зерновка пшеницы?
Строение семени однодольных растений рассмотрим на примере зерновки пшеницы (рис. 2.). У зерновки тонкий околоплодник срастается с семенной кожурой. Большую часть семени занимает мучнистый эндосперм, снизу к нему примыкает зародыш. Зародыш состоит из одной семядоли, которая называется щитком, зародышевы корешка, стебелька и почечки. Щиток непосредственно граничит с эндоспермом и осуществляет всасывание питательных веществ зародышем при прорастании семени.
Рис. 2. Строение семени пшеницы
Вопрос 5. Как расположен эндосперм у разных однодольных растений?
У других однодольных растений (лук, ландыш) эндосперм окружает зародыш со всех сторон. У пшеницы и других злаков эндосперм прилегает к зародышу с одной стороны. Среди однодольных имеются растения, семена которых не содержит эндосперма (чистяк). Семя у чистяка состоит из тонкой кожуры и зародыша, в котором сосредоточены все запасы, накопленные при созревании семени.
Вопрос 6. Чем различаются зародыши двудольных и однодольных растений?
У двудольных растений зародыш имеет две семядоли. Зародыш однодольных имеет только одну семядолю.
Питательные вещества у фасоли где обычно находятся
Фасоль – это двудольное растение, поэтому запас его питательных веществ откладывается сразу в двух семядолях и зародыше. Каковы особенности строения и где еще могут запасаться полезные вещества?
Немного истории
Родиной растения считают Америку, ведь именно там его начали культивировать. В Европе продукт оказался через некоторое время после открытий Колумба.
Изначально европейцы использовали культуру лишь в декоративных целях. Побеги растения красиво обвивали беседки, фасады зданий и частных домов. Затем ее стали выращивать и для употребления в пищу.
Только через 200 лет о фасоли узнали жители России. Они пошли вслед за европейцами, тоже использовали ее как украшение и декорации. Потом изучили положительное влияние растения на организм.
Как образуются семена фасоли?
Важный этап размножения растений – опыление. Именно при нем образуются семена фасоли. Пыльца содержит мужские половые клетки, попадает на зародыши с женскими. Мужская клетка достигает завязи, в течение 2-3 дней обычно формируется зародыш. Продолжительность цветения обычно составляет 2 недели. Спаржевые виды фасоли могут цвести гораздо дольше.
В плоде содержится 2-8 семян с оболочкой и запасом питательных веществ. Каждое зернышко – это зачаток жизни нового растения.
Каково строение фасоли?
Фасоль обычно имеет довольно крупные семена, которые хорошо прорастают. Семя растения считается органом для размножения. Также они выполняют функцию выживания и адаптации к изменениям окружающей среды.
Где в фасолевом семени находится запас питательных веществ?
Запас питательных веществ находится в клетках зародыша и семядолях. У некоторых видов запас располагается под кожурой рядом с зародышем будущего растения.
Чтобы увидеть зародыш, семена фасоли нужно предварительно размочить в воде, опустив их туда на 1-2 суток. Также можно поместить фасоль в мокрую ткань и оставить на столе. Затем с семечки нужно аккуратно снять кожу, надрезав ее, отделить друг от друга семядоли. Между ними можно будет найти зародыш.
Что нужно для прорастания семени?
Для прорастания новой жизни важно следующее:
- Умеренное поступление влаги (без избытка).
- Кислород, так как изначально зародыш поглощает его.
- Качественные, жизнеспособные семена.
- Достаточное освещение (некоторые сорта могут прорасти и без света).
- Температура выше 10 градусов, так как растение теплолюбиво.
- Довольно глубокая посадка (4-5 см). Если посадить глубже, то это может негативно сказаться на росте растения.
В чем состоят положительные свойства фасоли?
Культура обладает следующим влиянием на организм человека:
- Устранение проблем с опорожнением кишечника.
- Своевременный вывод шлаков и токсинов из организма.
- Снижение уровня вредного холестерина, укрепление сердечно-сосудистой системы.
- Снятие головной боли.
- Избавление от синдрома хронической усталости.
- Помощь при суставных заболеваниях.
- Улучшение иммунитета, помощь в лечении простудных заболеваний.
- Благотворное влияние на мозг и нервную систему.
- Укрепление костей, снижение риска возникновения остеопороза.
- Омоложение организма за счет улучшения состояния кожных покровов.
- Помощь при заболеваниях глаз.
Заключение
Таким образом, запас питательных веществ у растения находится в зародыше. Его защищает плотный слой кожуры, а также семядоли. Успешных рост культуры также зависит от многих внешних и внутренних факторов.
Обеспечение питательными веществами и рост растений
Глава
-
7
Цитаты -
381
Загрузки
Реферат
В качестве введения можно выделить две основные особенности питания растений. Во-первых, поскольку все физиологические процессы в растении в конечном итоге зависят от включения одного или нескольких минеральных питательных веществ в форме, соответствующей лежащей в основе биохимии, увеличение размеров органов растения и их правильное функционирование в конечном итоге зависят от соответствующей доступности необходимых питательные вещества [15, 16, 21].Во-вторых, степень, в которой процессы роста зависят от текущего потребления доступных извне питательных веществ, будет зависеть от количества и доступности хранимых питательных веществ и степени их рециркуляции внутри растения. Как хранение, так и рециркуляция — это явления, которые могут иметь решающее значение для выживания и приспособленности людей в условиях дефицита питательных веществ или в условиях колебания доступности питательных веществ. В отличие от проростков, выращенных из клеточной культуры, семена могут, например, содержать достаточное количество питательных веществ, чтобы обеспечить значительное увеличение размера растений без дальнейшего поглощения питательных веществ.У проростков, размножающихся на микроорганизмах, запас питательных веществ будет минимальным, и текущий рост, как ожидается, будет в значительной степени зависеть от текущей доступности и потребления минеральных питательных веществ. В первой части этой статьи обсуждаются некоторые основные взаимосвязи между поступлением питательных веществ и ростом растений. Подчеркивается возможность количественного контроля над продуктивностью сухого вещества путем регулирования подачи любого одного минерального питательного вещества. Во второй части обсуждаются качественные различия в ответной реакции роста в зависимости от характера ограничения питательных веществ.Предполагается, что, сочетая количественные и качественные аспекты, можно выращивать растения предсказуемого размера, морфологии, физиологии и биохимии. Считается, что это может иметь особое отношение не только к коммерческому приготовлению проростков заданного размера и внешнего вида, но также и к последующему производству и выделению определенных метаболитов из растений, микроразмножение которых с самого начала сочеталось с соответствующей практикой питания. .
Ключевые слова
Относительная скорость роста Конкретная площадь листа Минеральные питательные вещества Физиол Норма подачи растений
Эти ключевые слова были добавлены машиной, а не авторами.Это экспериментальный процесс, и ключевые слова могут обновляться по мере улучшения алгоритма обучения.
Это предварительный просмотр содержимого подписки,
войдите в
, чтобы проверить доступ.
Предварительный просмотр
Невозможно отобразить предварительный просмотр. Скачать превью PDF.
Ссылки
-
1.
Эдвардс Д.Г. и Ашер С.Дж. (1974) Значение скорости потока раствора в экспериментах с проточной культурой. Растения и почва 41: 161–175
CrossRefGoogle Scholar
-
2.
Ericsson T (1981) Рост и питание трех клонов Salix в растворах с низкой проводимостью. Physiol Plant 52: 239–244
CrossRefGoogle Scholar
-
3.
Ericsson T и Ingestad T (1988) Питание и рост проростков березы при различных относительных дозах добавления фосфора. Physiol Plant 72: 227–235
CrossRefGoogle Scholar
-
4.
Ericsson T и Kähr M (1993) Рост и питание проростков березы в зависимости от скорости поступления калия.Деревья 7: 78–85
CrossRefGoogle Scholar
-
5.
Ericsson T, Rytter L и Linder S (1992) Динамика питания и потребности лесов с коротким оборотом. В: С. П. Митчелл, Дж. Б. Форд-Робертсон, Т. Хинкли и Л. Сеннерби-Форсс, ред. Экофизиология лесных культур с коротким севооборотом, стр. 35–65. Лондон и Нью-Йорк: Elsevier Applied Science
Google Scholar
-
6.
Göransson A (1993) Рост и питание небольших
Betula pendula
растений, растущих при различных относительных дозах добавления железа.Деревья (в печати)
Google Scholar
-
7.
Göransson A (1993) Рост и питание мелких
Betula pendula
растений, растущих при различных относительных дозах добавления марганца. Tree Physiol (в печати)
Google Scholar
-
8.
Ingestad T (1981) Питание и рост проростков березы и ольхи серой в растворах с низкой проводимостью и при различных относительных скоростях добавления питательных веществ. Physiol Plant 52: 454–466
CrossRefGoogle Scholar
-
9.
Ingestad T (1982) Относительная скорость добавления и внешняя концентрация; управляющие переменные, используемые в исследованиях питания растений. Plant Cell Environ 5: 443–453
CrossRefGoogle Scholar
-
10.
Ingestad T и Kähr M (1985) Питание и рост хвойных саженцев при различной относительной скорости добавления азота. Physiol Plant 65: 109–116
CrossRefGoogle Scholar
-
11.
Ingestad T and Lund A-B (1979) Азотный стресс у проростков березы: I.Техника роста и рост. Physiol Plant 45: 137–148
CrossRefGoogle Scholar
-
12.
Ingestad T. и Lund A-B (1986) Теория и методы устойчивого минерального питания и роста растений. Scand J For Res 1: 439–453
CrossRefGoogle Scholar
-
13.
Ingestad T and McDonald AJS (1989) Взаимодействие между азотом и плотностью потока фотонов в проростках березы при стационарном питании. Physiol Plant 77: 1–11
CrossRefGoogle Scholar
-
14.
Marschner H (1986) Механизмы захвата ионов отдельными клетками и корнями: транспорт на короткие расстояния. В: Х. Маршнер, Минеральное питание высших растений, стр. 7–69. Лондон: Academic Press
Google Scholar
-
15.
Marschner H (1986) Функции минеральных питательных веществ: макроэлементы. В: Х. Маршнер, Минеральное питание высших растений, стр. 195–267. Лондон: Academic Press
Google Scholar
-
16.
Marschner H (1986) Функции минеральных питательных веществ: микроэлементы.В: Х. Маршнер, Минеральное питание высших растений, стр. 269–340. Лондон: Academic Press
Google Scholar
-
17.
McDonald AJS, Ericsson A and Lohammar T (1986) Зависимость запаса крахмала от доступности питательных веществ и плотности потока фотонов у мелкой березы (
Betula pendula
Roth). Plant Cell Environ 9: 433–438
CrossRefGoogle Scholar
-
18.
McDonald AJS, Lohammar T и Ingestad T (1992) Чистая скорость ассимиляции и развитие площади побегов у березы (Betula pendula Roth.) при различных установившихся значениях питания и плотности потока фотонов. Деревья 6: 1–6
CrossRefGoogle Scholar
-
19.
McDonald AJS, Palmer S и Davies WJ (1993) Растяжение листьев и корней при различной доступности нитратов у подсолнечника (Helianthus annuus L.). Physiol Plant (в печати)
Google Scholar
-
20.
Менгель К. и Киркби Е.А. (1987) Поглощение ионов и ионный статус растений. В: К. Менгель и Е. А. Киркби, Принципы питания растений, 4-е издание, стр. 113–147.Берн: Международный институт калийных удобрений
Google Scholar
-
21.
Менгель К. и Киркби Е.А. (1987) Питание и рост растений. В: K Mengel and EA Kirkby, Principles of Plant Nutrition, 4
th
edition, pp 247–302. Берн: Международный институт калийных удобрений
Google Scholar
-
22.
Olsen C (1950) Значение концентрации на скорость поглощения ионов высшими растениями в водных культурах. Physiol Plant 3: 152–164
CrossRefGoogle Scholar
-
23.
Pettersson R и McDonald AJS (1992) Влияние повышенной концентрации углекислого газа на фотосинтез и рост мелких растений березы (
Betula pendula
Roth.) При оптимальном питании. Plant Cell Environ 15: 911–919
CrossRefGoogle Scholar
-
24.
Терри Н. (1983) Ограничивающие факторы фотосинтеза. IV. Стресс-опосредованные железом изменения светособирательной и электронной способности и их влияние на фотосинтез
in vitro
.Plant Physiol 71: 855–860
PubMedCrossRefGoogle Scholar
Информация об авторских правах
© Springer Science + Business Media Dordrecht 1994
Авторы и аффилированные лица
- 1. Кафедра экологии и исследований окружающей среды Шведский университет сельскохозяйственных наук
0Uppsala 90, Швеция Взаимодействие с почвой: поглощение питательных веществ | Изучайте науку в Scitable
Бейер П. Золотой рис и
«Золотые» культуры для питания человека. Новое
Биотехнология 27 , 478-481 (2010).Бритто, Д. Т. и Х. Дж. Кронзукер.
Клеточные механизмы транспорта калия в растениях. Physiologia Plantarum 133 ,
637-650 (2008).Коннолли, Э. Л. и Э. Л. Уокер.
Пора качать железо: механизмы сигнализации о дефиците железа у высших растений. Current Opinion in Plant Biology 11 , 530-535 (2008).Фергюсон Б. Дж. и др. . Молекулярный анализ бобовых культур
Развитие узелков и ауторегуляция. Журнал
интегративной биологии растений 52 ,
61-76 (2010).Грэм Л., Грэм Дж. И
Л. Уилкокс. Биология растений. Река Аппер Сэдл, Нью-Джерси: Pearson Prentice Hall, 2006.Guerinot M. L. & Y. Yi.
Железо: питательно, ядовито и труднодоступно. Физиология растений 104 ,
815-820 (1994).Ад Р. и Х. Гиллебранд.
Концепции заводов по добыче и распределению полезных ископаемых. Текущее мнение в области биотехнологии
12 , 161-168 (2001).Джонс Б. и К. Юнг.
Подземное освоение космоса: развитие архитектуры корневой системы. Current Opinion in Plant Biology 15 , 97-102 (2012).Карандашов В. и М. Бухер.
Симбиотический транспорт фосфата у арбускулярных микориз. Тенденции в растениеводстве 10 ,
22-29 (2005).Лопес-Бусио Дж. и др. . Роль доступности питательных веществ
в регулировании корневой архитектуры. Текущий
Заключение по биологии растений 6 ,
280-287 (2003).Лимпенс Э. и Т. Бисселинг.
Сигнализация в симбиозе. Текущее мнение
в области биологии растений 6 , 343-350
(2003).Nehls U. и др. . Сахар для моего меда: углеводы
расщепление в эктомикоризном симбиозе. Фитохимия
68, , 82-91 (2007).Nehls U. Мастеринг
эктомикоризный симбиоз: влияние углеводов. Журнал экспериментальной ботаники 59 ,
1097-1108 (2008).Пио Ю. Дж. и др. . Высокое сродство K +
Транспортировка арабидопсиса: AtHAK5 и AKT1 имеют жизненно важное значение для создания рассады
и рост после прорастания в условиях низкого содержания калия. Физиология растений 153 ,
863-875 (2010).Sprent J. 60 млн лет зернобобовых культур
узловатость. Какие новости? Что меняется? Журнал
Экспериментальной ботаники 59 , г.
1081-1084 (2008).Вэнс К. Симбиотик
Азотфиксация и поглощение фосфора. Питание растений в мире
Снижение возобновляемых ресурсов. Завод
Физиология 127 , 390-397 (2001).Very, A. & H. Sentenac.
Молекулярные механизмы и регуляция транспорта K + в высших
Растения. Ежегодный обзор биологии растений 54 , 575-603 (2003).границ | Подача питательных веществ из окружающей среды напрямую меняет свойства растений, но косвенно определяет скорость роста вирусов
Введение
Экологическая стехиометрия (Sterner and Elser, 2002; Hessen et al., 2013) и конкуренция за ресурсы (MacArthur, 1972; Tilman, 1982; Miller et al., 2005) представляют собой две мощные теоретические основы для понимания воздействия измененных запасов питательных веществ на физиология и экология организмов.Обе концепции основываются на наблюдении, что виды различаются по своим требованиям к скорости подачи и соотношению различных элементарных питательных веществ. Экологическая стехиометрия в первую очередь основана на предположении, что стехиометрический баланс или соотношение нескольких химических элементов (например, углерода [C], азота [N] и фосфора [P]), доступных организмам, является движущей силой экологических процессов (Sterner и Elser, 2002; Hessen et al., 2013; Hillebrand et al., 2014). В частности, эта структура использовалась для изучения влияния соотношений C: N: P в ресурсе (например,g., добыча) на рост потребителей (Sterner, Elser, 2002; Hessen et al., 2013; Hillebrand et al., 2014). Напротив, теория ресурсной конкуренции предсказывает сохранение популяций, рост популяции и сосуществование видов на основе как скорости, так и соотношений питательных ресурсов и основана на предположении о конкурентных взаимодействиях между видами (например, потребителями и добычей) за общие ресурсы (MacArthur, 1972). ; Miller et al., 2005). Первоначально основанный на водных и морских экосистемах (Redfield, 1958; Corner et al., 1976; Tilman, 1976, 1977), экологическая стехиометрия и конкуренция за ресурсы использовались для оценки влияния добавления питательных веществ на численность, разнообразие и функциональные характеристики сосуществующих видов свободноживущих организмов в наземных экосистемах (Haddad et al., 2000 ; Cardinale et al., 2009; Zehnder, Hunter, 2009; Elser et al., 2010; Borer et al., 2014a, b; Seabloom et al., 2015a).
Помимо свободноживущих организмов, экологическая стехиометрия (Sterner and Elser, 2002) и конкуренция за ресурсы (Miller et al., 2005) также можно использовать для интерпретации влияния скорости и соотношения подачи питательных веществ на динамику микробов (Smith, 1993, 2007; Smith and Holt, 1996; Aalto et al., 2015). Обе теоретические основы предсказывают, что изменения в снабжении организма питательными веществами могут повлиять на размножение микроорганизмов (т. Е. Титр или размер популяции), поскольку содержание питательных веществ в организме хозяина может ограничивать производство требующих питательных веществ микробных клеток и частиц (Smith, 2007). В соответствии с этими прогнозами было показано, что скорость роста микроорганизмов человека и животных, вирусов водорослей и бактерий у ракообразных связана со стехиометрией питательных веществ (Elser et al., 2003; Карпинец и др., 2006; Класен и Эльзер, 2007; Frost et al., 2008; Lange et al., 2014; Маат и Брюссаард, 2016). Нормы и соотношения поступления питательных веществ также могут контролировать динамику различных инфекционных заболеваний растений и насекомых (Mitchell et al., 2003; Bedhomme et al., 2004; Borer et al., 2010, 2014b; Seabloom et al., 2013, 2015b; Lacroix et al., 2014). Однако независимо от того, влияют ли ресурсы окружающей среды на экологию вирусов растений на разных этапах цикла заражения и посредством прямого воздействия на рост вируса или косвенного воздействия на партнеров (т.е., завод-хозяин и конкуренты) интерактивной сети, ведущей к эпидемиям, остается неясным.
Применение стехиометрической теории и теории конкуренции за ресурсы к взаимодействиям хозяин-микроб может оказаться непростым делом, потому что запасы питательных веществ в окружающей среде и доступные для хозяев соотношения могут изменять динамику болезни на разных этапах эпидемиологического цикла (т. Е. Успешность заражения после инокуляции, при размножении хозяев, и передача между хозяевами; Aalto et al., 2015; Seabloom et al., 2015b; Борер и др., 2016). Кроме того, на каждую из этих стадий можно влиять различными путями (Borer et al., 2016), включая: (i) прямое влияние добавления питательных веществ к хозяевам на скорость и соотношение ограничивающих питательных веществ, доступных для репликации патогенов внутри хозяина. (Smith et al., 2005), (ii) косвенные эффекты доступности питательных веществ для очагового патогена, опосредованные взаимодействиями с другими конкурирующими патогенами (Smith and Holt, 1996; Smith, 2007; Lacroix et al., 2014; Lange et al., 2014), и (iii) косвенные эффекты снабжения питательными веществами, опосредованные изменениями скорости роста, размера и других функциональных характеристик хозяина (Whitaker et al., 2015).
Экологическая стехиометрия (Sterner and Elser, 2002) и теория конкуренции за ресурсы (Miller et al., 2005) предсказывают, что содержание и соотношения питательных веществ, доступных в организме хозяина, могут напрямую ограничивать производство молекул, таких как нуклеиновые кислоты и белки, которые необходимы для цикл заражения микроорганизмов. Облигатные паразиты, такие как вирусы растений, полностью полагаются на своего хозяина для завершения нескольких этапов своего эпидемиологического цикла, включая проникновение в хозяина и накопление внутри хозяина; а внутри хозяев вирусы захватывают молекулы хозяина, богатые азотом и фосфором, и метаболические пути (Maule et al., 2002; Стернер и Эльзер, 2002; Ahlquist et al., 2003; Elser et al., 2010). В контролируемых условиях скорость поступления азота и фосфора в растения-хозяева из окружающей среды, но не соотношение, увеличивает вероятность успешного установления инфекции (Bawden and Kassanis, 1950a; Lacroix et al., 2014; Smith, 2014). Скорость накопления внутри хозяина также может контролироваться ресурсами хозяина и стехиометрией питательных веществ хозяина (Spencer, 1941; Bawden and Kassanis, 1950b; Adam et al., 1987; Eraslan et al., 2007; Dordas, 2009; Alexander, 2010; Rua и другие., 2013). Кроме того, поскольку рост популяции патогенов внутри хозяина часто коррелирует со скоростью передачи новым хозяевам (Froissart et al., 2010), изменения в росте внутри хозяина, вызванные поставками питательных веществ в хозяине, могут изменить вторичные случаи передачи и динамику заболевания.
Скорости и соотношения питательных веществ в организме хозяина также могут косвенно влиять на рост патогенов внутри хозяина и передачу между хозяевами, влияя на сосуществование между коинфекционными патогенами из-за межвидовых вариаций стехиометрических требований C: N: P (Jover et al., 2014; Смит, 2014; Aalto et al., 2015), а также могут изменять взаимодействия между сообществом патогенов внутри хозяина. Растения могут являться хозяевами многих микроорганизмов (Seabloom et al., 2009; Roossinck, 2012), а межвидовые микробные взаимодействия внутри хозяина могут варьироваться от антагонистических до нейтральных и способствующих с различными потенциальными последствиями для динамики болезни (Turner, 2005; Rigaud et al., 2010; Elena et al., 2014; Seabloom et al., 2015b). Например, увеличение количества азота может снизить конкуренцию между вирусами и повысить уровень успешности инфекции и коинфекции (Lacroix et al., 2014). Конкуренция питательных веществ между микробами, разделяющими одного хозяина, также может изменить динамику заболевания, изменяя накопление в хозяине, вирулентность (т. Е. Пагубное влияние инфекции на приспособленность хозяина), скорость передачи и возникновение болезни (Smith and Holt, 1996; Al-Naimi et al., 2005; Pedersen, Fenton, 2007; Alizon et al., 2013; Hall, Little, 2013; Salvaudon et al., 2013; Lange et al., 2014; Borer et al., 2016).
Обеспечение питательными веществами окружающей среды также может косвенно влиять на динамику патогенов на различных этапах инфекционного цикла, изменяя функциональные характеристики хозяина.Например, было показано, что азот увеличивает концентрацию вируса растений за счет своего воздействия на биомассу хозяина, а не за счет прямого воздействия на вирус (Whitaker et al., 2015). Функциональные признаки, соответствующие морфологическим, физиологическим и фенологическим свойствам хозяина, могут в конечном итоге влиять на приспособленность организмов в различных условиях окружающей среды через эффекты роста, воспроизводства и выживания (Westoby, 1998; Westoby and Wright, 2006; Violle et al., 2007, 2014). ). Многие из этих черт отражают влияние истории эволюции, условий окружающей среды и компромиссов при распределении ограниченных ресурсов на каждый компонент приспособленности организма.В частности, различия в стратегии видов растений по приобретению, использованию и распределению питательных ресурсов были охарактеризованы на основе измерений набора взаимосвязанных функциональных признаков (Craine et al., 2002; Wright et al., 2004; Reich, 2014). В экономическом спектре «быстро-медленных» растений быстрорастущие растения обычно связаны с относительно низким соотношением C: P и N: P в тканях (то есть более высоким P) и повышенным выделением P-богатой рибосомной РНК (Elser et al., 2010). Листья быстрорастущих растений также имеют тенденцию быть недолговечными и структурно хрупкими, с тонкой пластинкой, низкой массой листьев на площадь (LMA), высокой фотосинтетической способностью и скоростью дыхания в темноте (Wright et al., 2004; Elser et al., 2010; Райх, 2014). Было показано, что межвидовые различия в среднем фенотипе растений в экономическом спектре «быстро-медленно» влияют на способность различных видов растений действовать в качестве эффективных резервуаров вирусов растений (Cronin et al., 2010) и внутривидовых Изменение функциональных характеристик растений в ответ на поступление питательных веществ в окружающую среду также может изменить эпидемиологические параметры (Whitaker et al., 2015).
Микробная инфекция также может изменять фенотип хозяина, повышая возможность обратной связи между поступлением питательных веществ и инфекцией патогенов на свойства растений.Облигативных паразитов, таких как вирусы, можно рассматривать как потребителей (Aalto et al., 2015), которые конкурируют со своим хозяином за питательные ресурсы, что может привести к повышению вирулентности при истощении ресурсов хозяина (Smith and Holt, 1996; Smith, 2007). В этом случае вирулентность патогенов может развиваться за счет компромисса, если выгоды от увеличения репликации внутри хозяина и коррелированного увеличения передачи между хозяином происходят за счет увеличения пагубного воздействия на приспособленность хозяина за счет эксплуатации ресурсов хозяина (Alizon et al. al., 2009, 2013; Froissart et al., 2010; Doumayrou et al., 2013). В конечном итоге эпидемиология горизонтально передаваемых патогенов может быть изменена этими эффектами вирулентности на рост и продолжительность жизни хозяина, что может быть аппроксимировано несколькими характеристиками, связанными со спектром экономических характеристик растения (например, LMA, скоростью роста, продолжительностью жизни листьев) и этой обратной связью. может изменить взаимодействие между скоростью поступления питательных веществ в растения и ростом и распространением патогенов растений.
В целом, экологическая стехиометрия (Sterner and Elser, 2002) и конкуренция за ресурсы (Miller et al., 2005) теория предсказывает, что запасы питательных веществ в организме хозяина могут влиять на экологию патогенов, таких как вирусы растений, на различных стадиях их жизненного цикла за счет прямого воздействия скорости или соотношения доступных питательных веществ в организме хозяина. На каждую стадию эпидемиологического цикла также могут влиять изменения в динамике коинфекционных патогенов и функциональных характеристик хозяина, опосредованных питательными веществами хозяина. Здесь мы экспериментально проверили влияние скорости поступления азота и фосфора, содержания питательных веществ в растениях и функциональных характеристик, а также наличия сопутствующего инфицирующего микроба на накопление внутри хозяина, вирулентность и передачу между хозяином двух видов вирусов растений, ячменя. вирус желтого карлика-PAV (BYDV-PAV) и вирус желтого карлика злаков-RPV (CYDV-RPV).Мы использовали факторную комбинацию двух скоростей подачи питательных веществ N и P, что позволило создать четыре обработки питательными веществами со стехиометрическими соотношениями, воспроизводимыми при низких и высоких скоростях подачи питательных веществ. Измеряя содержание углерода и питательных веществ в растениях, а также стехиометрию C: N: P, мы смогли проверить, зависят ли процессы в первую очередь от содержания растительной ткани или соотношения питательных веществ. Мы проверили роль конкурента внутри хозяина в динамике заражения, включив в него однократно инфицированных и совместно инфицированных хозяев. Нашим центральным хозяином был Avena sativa ( Poaceae ), широко культивируемый хозяин этой группы вирусов.Мы использовали этот дизайн, чтобы ответить на следующие вопросы:
• Меняют ли запасы питательных веществ и инфекция стехиометрии и свойств растений?
• Могут ли запасы питательных веществ и стехиометрия тканей хозяина предсказать титр вируса внутри хозяина?
• Каково относительное значение снабжения растений питательными веществами, стехиометрии и свойств растений, а также коинфекции для накопления вируса внутри хозяина и передачи вируса между хозяином?
Материалы и методы
Учебная система
Вирусы желтых карликов ячменя и злаков (B / CYDVs, Luteoviridae) являются универсальными хозяевами и, как известно, инфицируют не менее 150 видов трав семейства Poaceae (Irwin and Thresh, 1990; D’arcy and Burnett, 1995).Инфекция у растений носит системный характер, но ограничивается клетками флоэмы-хозяина. Заражение растений B / CYDV может быть связано с проявлением различных симптомов, включая карликовость, пожелтение и покраснение, а также с серьезными потерями урожая (Irwin and Thresh, 1990; Perry et al., 2000). B / CYDV также могут изменять различные характеристики растений, такие как плодовитость и долголетие хозяев, и были признаны предшественниками резкого изменения видового состава растений на естественных лугах Калифорнии (Malmstrom et al., 2005; Борер и др., 2007).
Группа B / CYDV распространена по всему миру и включает представителей родов лютеовируса (например, BYDV-PAV) и полеровируса (например, CYDV-RPV), двух распространенных видов вируса B / CYDV, обнаруженных в обеих культурах. и дикие растения (Leclercq-Le Quillec et al., 2000; Robertson and French, 2007; Seabloom et al., 2010). Эти вирусы обязательно передаются от растения к растению через тлей-переносчиков (Aphididae) устойчивым, циркулирующим и нераспространяющимся образом (Miller and Rasochova, 1997; Gray and Gildow, 2003).По крайней мере 25 видов тлей известны как векторы B / CYDV, и эффективность передачи каждого вида вируса сильно различается среди переносчиков (Halbert and Voegtlin, 1995; Power and Gray, 1995; Miller and Rasochova, 1997). Тля вида Rhopalosiphum padi является эффективным переносчиком как BYDV-PAV, так и CYDV-RPV, основных вирусов в этом исследовании.
Изоляты B / CYDV и переносчики тли
Мы использовали по одному изоляту каждого из двух видов вируса, BYDV-PAV и CYDV-RPV, которые были первоначально собраны с зерновых культур в штате Нью-Йорк и сохранены в Dr.Лаборатория Стюарта Грея (Корнельский университет, США). В нашей лаборатории мы поддерживали эти изоляты, инокулируя новые культуры здоровых 10-дневных животных A. sativa cv. Костный черный овес (Poaceae; Национальная система зародышевой плазмы растений, USDA; США; далее A. sativa ) растения-хозяева, высаженные в горшки размером 15 × 15 см, содержащие горшечную почву Sunshine MVP (Sun Gro Horticulture, Массачусетс, США), каждые 3 недели после инокуляции. процедура описана ниже.
Невирулевые тли R. padi выращивали в горшках размером 15 × 15 см, каждый из которых был засеян 15 здоровыми A.sativa в горшечной почве Sunshine MVP (Sun Gro Horticulture, Массачусетс, США). Колонии поддерживали в отдельной камере для выращивания (27 ° C, 16 часов дня, 8 часов ночи, флуоресцентные лампы мощностью 32 Вт) и поливали дважды в неделю 300 мл водопроводной воды. Примерно 100 тлей переносили каждые 2 недели на здоровые 10-дневные растения A. sativa .
Рост растения-хозяина
Семена A. sativa высевали в горшки диаметром 3,8 см и глубиной 21 см, в горшки объемом 164 мл, содержащие водонасыщенную смесь 70/30% (об. / Об.) Sunshine, высшего сорта, средний вермикулит (Sun Gro Horticulture, Массачусетс). , США) и заливочный материал Turface MVP (Turface Athletics, Иллинойс, США).Затем горшки помещали в контролируемые условия в камеру для выращивания, свободную от вирусов и тлей (23 ° C, 15 часов дня, 9 часов ночи, натриевые лампы высокого давления мощностью 400 Вт). Семенам давали 10-дневный период прорастания, в течение которого проростки прореживали до одного растения на горшок.
Экспериментальный дизайн
Растения случайным образом были разделены на четыре группы, которые были имитированы, индивидуально или совместно инокулированы BYDV-PAV или CYDV-RPV. В каждой группе мы инокулировали по семь растений на одну обработку удобрением, различающихся по скорости поступления азота и фосфора (Ctrl [7.5 мкМ, 1 мкМ]; N [375 мкМ, 1 мкМ]; P [7,5 мкМ, 50 мкМ] и NP [375 мкМ, 50 мкМ]; соответственно; Таблица S1). Вся процедура повторялась трижды. Таким образом, в каждом из наших 16 экспериментальных условий (4 типа прививок [Mock, BYDV-PAV, CYDV-RPV, BYDV-PAV + CYDV-RPV]) * (4 обработки питательными веществами [Ctrl, N, P, NP]), и из-за потери нескольких растений у нас было от 19 до 21 растения.
Обработка удобрением представляет собой полную факторную комбинацию двух уровней добавления азота и фосфора в концентрациях, эквивалентных 0.2 и 10% питательного раствора Хогланда половинной концентрации (Hoagland and Arnon, 1938; Downs and Hellmers, 1975), диапазон скорости подачи питательных веществ, известный как изменение успешности вирусной инфекции в этой системе (Lacroix et al., 2014), в то время как концентрации других макро- и микроэлементов оставались постоянными (Таблица S1). Каждое растение дважды в неделю удобряли 30 мл питательного раствора.
Мнимая вакцинация и вирусные прививки
Мы провели инокуляцию растений, когда у них было два листа, в соответствии с ранее опубликованным протоколом (Lacroix et al., 2014) с изменениями из (Gray, 2008). Вкратце, после 2-часового периода голодания невирулезным тлям давали 48-часовой период доступа к вирусу на листьях, которые были отделены либо от неинфицированных растений, либо от хозяев, индивидуально инфицированных BYDV-PAV или CYDV-RPV и помещенных в вертикальные стеклянные флаконы 12 × 1,5 см в камере для выращивания (23 ° C, 16 часов дня, 8 часов ночи, люминесцентные лампы мощностью 40 Вт). После этого периода приобретения тлей, которые питались отслоившимися листьями одного и того же типа инфекции (не инфицированные BYDV-PAV и CYDV-RPV), объединяли вместе.После еще двухчасового голодания мы перенесли по пять тлей на каждое имитационное и индивидуально инокулированное растение и по десять особей на каждое совместно инокулированное растение (по пять особей из каждой партии тлей, которые питались BYDV-PAV и CYDV-RPV). тканевый материал). Тлю помещали в клетку из полиэстера размером 8 × 2,5 см и 118 мкм (Sefar America Incorporated, Канзас-Сити, штат Миссури, США), прикрепленную к самому молодому из возможных листьев каждого экспериментального растения (возрастом 10 дней). Затем растения помещали в камеру для выращивания (23 ° C, 16 часов дня, 8 часов ночи, флуоресцентные лампы мощностью 40 Вт) и тлей убивали с использованием инсектицидного мыла (Ortho) после 72-часового периода доступа для инокуляции вируса.
Обнаружение вирусов с помощью ОТ-ПЦР
Через 19 дней после инокуляции (dpi), то есть в момент времени, когда вирусная инфекция стала системной и титр вируса превышает порог обнаружения (Chain et al., 2005), был собран 20-сантиметровый кусок первого листа каждого растения. и хранили при -20 ° C для дальнейшего обнаружения вирусов. Статус заражения каждого тестируемого растения проверяли, как описано Lacroix et al. (2014). Вкратце, экстракцию тотальной РНК проводили с использованием Trizol (Invitrogen, Grand Island, NY) в соответствии с инструкциями производителя.Затем экстракты РНК хранили при -20 ° C до использования. Чтобы оценить статус заражения каждого растения, мы использовали мультиплексный анализ RT-PCR, который дает фрагменты разного размера для BYDV-PAV (т.е. 298 п.н.) и CYDV-RPV (т.е. 447 п.н.) с использованием специфических праймеров для BYDV-PAV (PAVR1, ATTGTGAAGGAATTAATGTA; PAVL1, AGAGGAGGGGCAAATCCTGT) и CYDV-RPV (RPVR2 CTGCGTTCTGACAGCAGG, RPV L ATGTTGTACCGCTTGATCCAC). Эти праймеры были адаптированы из ранее опубликованного протокола (Deb and Anderson, 2008). Продукты ПЦР визуализировали на 2% (мас. / Об.) Геле агарозы-1000 (Invitrogen), окрашенном SybrSafe (Invitrogen), с использованием УФ-системы EZ doc (Bio-Rad), и размер фрагмента проверяли сравнительно с лестницей ДНК 100 п.н. (Продукты Apex Bioresearch).
Титр вируса
Накопление каждого вида вируса внутри хозяина определяли на основе тех же экстрактов РНК, полученных, как описано выше, и с использованием протокола количественной ПЦР в реальном времени. Абсолютную количественную оценку титра вируса проводили на основе стандартной кривой, построенной для каждого вида вируса из 10-кратных серийных разведений транскриптов РНК известной концентрации. Эти транскрипты были получены с использованием набора MEGAscript ® T7 (Life Technologies) и ампликонов, полученных с помощью обычной ОТ-ПЦР со специфическими праймерами (как описано выше).Затем транскрипты очищали с использованием протокола осаждения фенолом / хлороформом и изопропанолом, как указано в инструкциях производителя. Размер полученных транскриптов проверяли на обычном 2% агарозном геле относительно лестницы ДНК 100 п.н. (Apex Bioresearch Products). Концентрацию транскриптов РНК (нг / мкл) определяли с помощью спектрофотометра с нанокапельками (Thermo Scientific). Полученное значение преобразовывали в моль / мкл, используя молекулярную массу рибонуклеотида (340 г / моль) и количество оснований каждого типа транскрипта (Nb).Применялась следующая математическая формула: концентрация РНК в моль / мкл = концентрация РНК в нг / мкл * (10 -9 нг / 1 г) * (1 моль / 340 г) * (1 / Nb). Константу Авогадро (6,023 × 10 23 молекул / моль) использовали для оценки количества копий транскрипта РНК на мкл.
Реакции RT-PCR
TaqMan ® были выполнены в конечном объеме 25 мкл с использованием наборов реагентов Master Mix для одностадийной RT-PCR (RNA-to-Ct 1-Step Kit, Applied Biosystems) и Applied Biosystems One Plus Real- система ПЦР времени.Были включены три повтора каждого серийного разведения транскриптов РНК, нематричного контроля (т.е. стерильная вода вместо РНК), контроля без амплификации (т.е. отсутствие ферментов в реакции) и экстракта РНК каждого тестового образца. в каждом пробеге. Реакции проводили с 2,5 мкл образца (т.е. экстрактов РНК или стерильной воды) в соответствии с инструкциями производителя. Обратную транскрипцию проводили в течение 30-минутного цикла при 48 ° C. После 15-минутного периода активации полимеразы Taq при 95 ° C фрагменты кДНК были амплифицированы после 45 циклов денатурации (25 с 95 ° C), отжига и удлинения (1 мин при 60 ° C).Амплификацию кДНК проводили с использованием специфических праймеров и зондов, прикрепленных к гасителю связывания малой бороздки (MGB) и к другому репортерному флуоресцентному красителю для BYDV-PAV (прямой праймер, TGGTCGCCCAAAAAATCTAAAAC; обратный праймер GGAGTAAGGCTCGCAGTAAATTGCCGCATAGACCCAC и зонд AGCCAGCAGCAC и зонд RPV (прямой праймер, GAGGTTAGCGAGGAGTTAGAATTC; обратный праймер, AACTACCTCAGAGTTGCCACATTC; и зонд, ACATCTTCAAGACTCCTAACCTCGCCAT, VIC).
Стандартные кривые были построены с использованием известной концентрации транскриптов РНК (log 10 геномных копий в 2.5 мкл) для каждого серийного разведения и соответствующий порог цикла (то есть Ct, количество циклов амплификации, необходимых для значительного увеличения флуоресценции репортера). Количественное определение РНК (log 10 геномных копий в 2,5 мкл) для тестируемых образцов рассчитывали на основе порогового значения Ct, полученного для каждого образца, и стандартных кривых. Затем титр вируса выражали как log 10 геномных копий на мг свежей ткани.
Характеристики растений
Растения оценивались по нескольким функциональным признакам, которые обычно измеряются для описания стратегии приобретения и использования ресурсов видов растений по всему спектру экономики растений (Craine et al., 2002; Райт и др., 2004; Райх, 2014). Измеряемые характеристики растений включают количество дней от ложной инокуляции или вирусной инокуляции до появления третьего листа (см. NbDaysEmerg), среднее содержание хлорофилла во втором листе по трем значениям на лист, измеренное с помощью SPAD-502 (15 dpi) ( Konica Minolta), процент площади стареющих (т.е. сухих) листьев, усредненный по первому и второму листу (например, Senes) 18 dpi, свежей биомассе (g) над (AG) и под землей (BG) и соотношение между этими двумя значениями (ABG) 41 dpi, сухой массой листа на единицу площади (LMA, мг · см −2 ) и содержанием воды, измеренным как разница между свежей и сухой массой листьев на площади (мг · см −2 ) 41 dpi.Мы также записали средний процент площади листьев на двух листьях, которые были покрыты хлоротическими симптомами, характерными для инфекции B / CYDV (то есть пожелтением и покраснением), 19 точек на дюйм.
Данные химического состава ткани, то есть содержание фосфора (P), азота (N) и углерода (C), были получены на основе надземной ткани листа, собранной с разрешением 41 dpi. Для каждого образца ткань листа сушили в печи при 65 ° C в течение 48 ч, а затем измельчали с помощью Beadbeater (Biospec). Для определения содержания N и C 3 мг сухой и измельченной ткани из каждого образца взвешивали в оловянной капсуле.Абсолютное количественное определение N и C проводили с использованием элементного (C: H: N) анализатора. Для анализа содержания P в тканях были приготовлены две копии по 1–1,5 мг каждая в оловянных капсулах. Количественное определение P проводили с использованием протокола разложения серной кислотой. Стандартная кривая была построена на основе данных, полученных для образцов яблочного стандарта NIST различной массы (от 0 до 6,9 мг) и известного содержания фосфора (т.е. 0,159% от веса сухой ткани). Все образцы в оловянных капсулах помещали в предварительно промытые кислотой и взвешенные стеклянные флаконы, а затем помещали в муфельную печь на 30 мин при 300 ° C, а затем на 2 ч при 550 ° C.После этого в каждый флакон добавляли 0,4 мл серной кислоты (10 N H 2 SO 4 , 10,8 M) и 5 мл наночистой воды. Затем закрытые флаконы автоклавировали в течение 30 минут при 121 ° C. Затем 1 мл молибдатного реагента комнатной температуры (т.е. из раствора, приготовленного из 0,208 г сурьмы-калия — тартрата, смешанного с 9,6 г гептамолибдата аммония 4-гидрата в 1 л воды с наночастицами) добавляли в стеклянные флаконы с комнатной температурой. Затем в каждый флакон добавляли 0,4 мл аскорбиновой кислоты (2 г / мл) и 3,2 мл воды Nanopure, чтобы довести конечный объем до 10 мл.Каждую пробирку взвешивали, и затем каждый образец анализировали при 880 нм с использованием кювет диаметром 1 см и спектрофотометра Varian. Содержание в тканях N, P и C выражали в молях, на основании чего рассчитывали соотношения питательных веществ в тканях (N: P, C: N, C: P).
Скорость передачи
Свежая ткань листьев, собранная с экспериментальных растений (т.е. исходных растений), 19 точек на дюйм использовалась для оценки скорости передачи. В соответствии с протоколом инокуляции, описанным выше, невирулезным тлям был предоставлен 48-часовой период доступа к ткани листа каждого исходного растения отдельно.После 2-часового периода голодания 5 тлей переносили в сетчатую клетку, прикрепленную к самому молодому из возможных листьев каждого из семи 10-дневных растений-реципиентов A. sativa на исходное растение. Растения помещали в камеру для выращивания (23 ° C, 16 часов дня, 8 часов ночи, флуоресцентные лампы мощностью 40 Вт) и тлей убивали с использованием инсектицидного мыла (Орто) после 72-часового периода доступа для инокуляции вируса. Растения-реципиенты удобряли дважды в неделю 30 мл питательного раствора Ctrl (7,5 мкМ N, 1 мкМ P). Ткань листа собирали с разрешением 19 точек на дюйм для оценки статуса инфекции каждого растения-реципиента с использованием протокола RT-PCR, как описано выше.Для каждого вида вируса скорость передачи выражалась как доля инфицированных растений-реципиентов на исходное растение.
Статистический анализ
Все статистические анализы были выполнены с использованием R версии 3.3.2 (R Foundation for Statistical Computing, Вена, Австрия).
Данные о признаках (т.е. необработанные значения всех признаков, за исключением симптомов, вызванных вирусом) всех экспериментальных растений были проанализированы с использованием анализа главных компонентов (PCA) из пакета FactoMineR. Перед анализом все переменные признаков растений были масштабированы до единичной дисперсии.Корреляции между переменными признака растения и размерами PCA определялись на основе вклада каждой переменной в каждое измерение и на основе результатов (то есть коэффициента корреляции и теста значимости), полученных с помощью функции dimdesc. Основные компоненты (т.е. ортогональные размеры, N = 5), которые в совокупности объясняют 80% дисперсии, включенной в исходный набор данных, были сохранены для дальнейшего анализа.
Чтобы оценить влияние оплодотворения и инфекции на характеристики растений, мы проанализировали влияние скорости поступления азота и фосфора в растение-хозяин, а также однократного и однократного поступления азота и фосфора.коинфекция по координатам отдельных растений по каждому из пяти сохраненных параметров PCA с использованием усреднения модели (Grueber et al., 2011). Этот подход позволил нам принять во внимание, что объясняющие переменные могут быть ковариантными и что может существовать более одной подходящей модели. Все переменные были стандартизированы перед анализом с использованием функции стандартизации в пакете arm R. Мы использовали функцию земснаряда в пакете MuMIn R, чтобы соответствовать всем возможным моделям. С помощью модели мы оценили значения параметров, ошибки и взвешенную важность AIC.avg в пакете MuMIn R. «Относительная важность» каждой независимой переменной была оценена на основе релятивизированной суммы весов Акаике, суммированных по всем моделям, в которых появляется параметр, которые находятся в пределах четырех единиц AIC C (т. Е. AIC с поправкой на небольшой размер выборки). модели с самым низким AIC. Значимость варьируется от 0 (параметр не имеет пояснительного веса) до 1 (параметр во всех топ-моделях).
Для каждого вида вируса различия в накоплении внутри хозяина в инфицированных растениях оценивали как функцию от скорости поступления азота и фосфора в растение-хозяин, от присутствия внутри хозяина по сравнению сотсутствие второго вида вируса (т. е. коинфекция) и признаки растений, представленные координатами отдельных растений по каждому из пяти сохраненных параметров PCA. Мы повторили этот анализ титра внутри хозяина как функции скорости подачи питательных веществ, коинфекции и исходных значений каждого измеренного признака растения вместо координат PCA. Переменная, соответствующая содержанию воды, была удалена из этого последнего анализа из-за внутренней корреляции с LMA. Мы использовали метод усреднения модели, как описано выше для этих двух типов анализа.Мы также применили линейную модель для оценки количества индуцированных вирусом симптомов, которая была преобразована log 10 для нормализации его распределения в зависимости от накопления вируса внутри хозяина.
Наконец, мы оценили различия в скорости передачи каждого вида вируса в отдельности в зависимости от скорости поступления питательных веществ в растение-хозяин, коинфекции и характеристик растения (т.е. представленных либо координатами отдельных растений по каждому из пяти сохраненных параметров PCA, либо исходными показателями каждого признака растения, кроме воды) с использованием модели усреднения.
Результаты
Характеристики растений
Первые пять основных компонентов, взятые вместе, объяснили 80,5% фенотипических вариаций, включенных в исходный набор данных (Таблица 1). Вместе первые два основных компонента объясняют 53,7% этой фенотипической вариации (Таблица 1 и Рисунок 1A). Главный компонент 1 был связан с листовой Р и признаками роста над землей. Образцы с более высокими значениями основного компонента 1 (т.е. Dim.1) соответствовали растениям с более низким содержанием фосфора, более высоким соотношением C: P и N: P листвы, более быстрым появлением листьев, более высоким содержанием хлорофилла, менее стареющей тканью, более высокой надземной биомассой. , а также соотношение надземной / подземной биомассы и более высокое содержание воды (Таблица 1 и Рисунок 1A).
Таблица 1 . Результаты анализа главных компонентов, выполненного со значениями признаков, измеренными на неинфицированных растениях и растениях, однократно или совместно инфицированных BYDV-PAV и / или CYDV-RPV.
Рисунок 1 . Координаты исходных переменных (например, признаки растений, показанные стрелками) и средние координаты не- (перечеркнутые квадраты), одиночного CYDV-RPV (белые кружки), одиночного BYDV-PAV (черные кружки) и ко- (серые треугольники) ) зараженные растения по основным компонентам (Разм.) 1 и 2 (A) и 4 и 5 (B) . Измеренные признаки растений сокращены, как в таблице 1. Вектор переменных, которые больше всего повлияли на Dim. 1, 2, 4 и 5 выделены жирным шрифтом. Планки погрешностей представляют ± 1 SEM.
Характеристики листьев, включая химию тканей и продукцию биомассы, различались по размерам 2 и 3 PCA. У растений с более высокими координатами по параметру 2 PCA было более низкое содержание азота в тканях и соотношение N: P, но более высокое соотношение C: N в тканях, LMA и содержание воды. (Таблица 1 и Рисунок 1A).Параметр 3 PCA объясняет 10,7% фенотипической дисперсии и контрастирует с высоким содержанием C: P в тканях листа с высокой подземной биомассой (Таблица 1). Измерения PCA 4 и 5 вместе объяснили 16,1% фенотипической изменчивости (Таблица 1 и Рисунок 1B). Более высокие значения по этим параметрам соответствовали отдельным растениям с более высокими значениями LMA (см. Параметр PCA 4), более высокой подземной биомассой и более низким соотношением наземной / подземной биомассы (см. Параметр PCA 5; Таблица 1 и Рисунок 1B). Соотношения питательных веществ были в значительной степени связаны с первыми тремя осями PC, но признак растения, наиболее сильно коррелировавший с каждым измерением PCA, был надземной биомассой (AG), содержанием азота в листьях (N), подземной биомассой (BG), LMA и соотношение надземной и подземной биомассы (ABG) соответственно (Таблица 1).
Добавление азота в окружающую среду и заражение измененных свойств растений
Повышенная скорость подачи азота к растениям-хозяевам увеличивалась ( p <0,001, относительная важность = 1) для отдельных растений по параметру PCA 1 (например, более высокий AG), но снижалась ( p = 0,006, относительная важность = 1). размер 4 (т.е. нижний LMA; Таблицы 2, 3). Увеличение скорости подачи фосфора не привело к значительному изменению значений отдельных растений по любому параметру PCA (таблицы 2, 3).
Таблица 2 . Краткое изложение эффектов скорости поступления азота и фосфора, однократного заражения BYDV-PAV или CYDV-RPV и совместного инфицирования на характеристики растений (т.е. координаты отдельных растений по основным компонентам [Разм.] 1 и 2) после усреднения модели.
Таблица 3 . Сводная информация о влиянии скорости поступления азота и фосфора, однократного заражения BYDV-PAV или CYDV-RPV и сочетанного инфицирования на характеристики растений (т.е. координаты отдельных растений по основному компоненту [Dim.] 3, 4 и 5) после усреднения модели.
Единичное инфицирование, будь то BYDV-PAV или CYDV-RPV, не повлияло значительно на индивидуальные показатели растений по параметрам PCA 1, 2 или 3 по сравнению с имитацией инокулированных растений (таблицы 2, 3 и рисунок 1A). Однако коинфекция снизила рост растений и изменила химический состав растений, т. Е. Коинфекция уменьшила координаты по параметру PCA 1 ( p = 0,01, относительная важность = 1, например, более низкие AG и C: P) и 2 ( p = 0,042, относительная важность = 0.23, например, более высокий N) по сравнению с имитацией инокулированных растений (таблица 2 и рисунок 1A).
Инфекция CYDV-RPV приводила к более толстым (т. Е. С более высоким LMA) листьям растений, то есть к увеличению значений растений по параметру PCA 4 ( p = 0,004, относительная важность = 0,1), и коинфекция не изменила эту взаимосвязь (Таблица 3 и рисунок 1B). Кроме того, растения, инфицированные CYDV-RPV, характеризовались более высоким соотношением надземной и подземной биомассы, то есть ассоциировались со значительно более низкими координатами по параметру PCA 5 ( p = 0.011, относительная важность = 1). Однако присутствие BYDV-PAV увеличивало координаты инфицированных CYDV-RPV хозяев ( p = 0,011, относительная важность = 1; Таблица 3 и Рисунок 1B) по параметру PCA 5. Признаки растений, инфицированных BYDV-PAV, не отличались от имитация инокулированных растений размером 4 и 5 РСА (таблица 3 и рисунок 1B).
CYDV-RPV, но не BYDV-PAV, накопление внутри хозяина коррелировало с изменениями в признаках растений
Повышенные нормы поступления азота и фосфора не повлияли напрямую на накопление вируса CYDV-RPV внутри хозяина (таблицы 4, 5).Однако титр CYDV-RPV был увеличен у растений с более высокими координатами PCA, размерностью 2 (например, высшее растение C: N и LMA, p = 0,039, относительная важность = 0,81, Таблица 4 и Рисунок 2A). Подробный анализ накопления вируса CYDV-RPV внутри хозяина как функции скорости поступления питательных веществ, коинфекции и значений каждого признака растения выявил положительную корреляцию с LMA ( p = 0,001, относительная важность = 1; Таблица 5 и Рисунок 2B ) и отрицательный эффект коинфекции ( p = 0.025, относительная важность = 0,91; Таблица 5 и Рисунок 3). На титр BYDV-PAV не влияли показатели поступления в окружающую среду, коинфекция или характеристики растений (таблицы 4, 5 и рисунок 3).
Таблица 4 . Краткое изложение эффектов скорости поступления азота и фосфора, коинфекции и характеристик растений (т.е. координаты отдельных растений по каждому главному компоненту [размер]) на титр CYDV-RPV и BYDV-PAV после усреднения модели.
Таблица 5 . Сводная информация о влиянии скорости поступления азота и фосфора, коинфекции и индивидуальных свойств растений на титры CYDV-RPV и BYDV-PAV после усреднения модели.
Рисунок 2 . Накопление CYDV-RPV внутри хозяина, измеренное на основе ткани листа с разрешением 19 dpi, как функция координат отдельных растений на втором главном компоненте (A) и значений LMA отдельных растений (B) . R-квадрат, p -значение и наклон показаны для значимых линейных двумерных отношений.
Рисунок 3 . Среднее накопление вируса в ткани листа 19 dpi для CYDV-RPV (белые кружки) и BYDV-PAV (черные кружки) при однократном и совместном заражении.Планки погрешностей представляют ± 1 SEM.
Накопление CYDV-RPV внутри хозяина коррелировало с хлоротическими симптомами
Процент площади инфицированного листа, который был обесцвечен (т.е. пожелтел и покраснел), положительно коррелировал с титром внутри хозяина для CYDV-RPV ( p = 0,004, рис. 4A), но не для BYDV-PAV ( p = 0,95, рис. 4С). Наконец, титр внутри хозяина не влиял напрямую на скорость передачи вирусов CYDV-RPV и BYDV-PAV (таблицы S2, S3 и рисунки 4B, D).Кроме того, на скорость передачи вируса существенно не влияли скорость поступления питательных веществ, химический состав тканей, коинфекция вируса или характеристики растений (таблицы S2, S3).
Рисунок 4 . Процент площади листьев с симптомами, ассоциированными с инфекцией вируса папилломы человека (A), и вирусом папилломы человека (C), и средняя доля растений, инфицированных после вторичной передачи от дифференцированно оплодотворенных опытных растений, в зависимости от вируса папилломы человека (B), и PAV (D). ) Накопление внутри хозяина, измеренное на основе ткани листа, 19 точек на дюйм.Титр, симптомы и скорость передачи вируса показаны для «исходных» одно- (белые кружки), однократно- (черные кружки) и ко- (серые треугольники) растений, инфицированных вирусом вируса папилломы. R-квадрат, значение p и линия показаны для значимых линейных двумерных отношений. Содержание азота и фосфора в растворах для удобрений Ctrl (контроль), N (азот), P (фосфор) и NP (азот и фосфор) составляет: 7,5 / 1, 375/1, 7,5 / 50, 375/50 мкМ, соответственно. Планки погрешностей представляют ± 1 SEM.
Обсуждение
Экология вирусов растений может находиться под влиянием множества абиотических и биотических факторов на различных стадиях их эпидемиологического цикла. Пути, с помощью которых питательные вещества влияют на рост и передачу вирусной популяции, могут включать как прямые эффекты увеличения запасов и содержания питательных веществ в организме хозяина на успех и рост вирусной инфекции, так и косвенные эффекты добавления питательных веществ через изменение динамики коинфекционных патогенов и фенотипа хозяина. В отличие от прогнозов экологической стехиометрии (Sterner and Elser, 2002) и конкуренции за ресурсы (Miller et al., 2005), мы не обнаружили какого-либо прямого влияния скорости и соотношения питательных веществ в источниках окружающей среды или растительной ткани на динамику вирусов в нашей системе. Повышенное снабжение азотом сместило фенотип растений в сторону более высокой скорости роста растений и более тонких листьев (то есть, уменьшило LMA), в соответствии с ожидаемыми эффектами азотных удобрений (Elser et al., 2007; Dordas, 2009; Reich, 2014), в то время как заражение CYDV- RPV приводил к более толстым листьям растений (т. Е. К более высокому LMA). Титр CYDV-RPV был выше у растений с более высоким LMA.Титр CYDV-RPV также снижался из-за присутствия BYDV-PAV, что дополнительно указывает на конкуренцию внутри хозяина между этими близкородственными вирусами (Lacroix et al., 2014). Симптомы хлоротии усиливаются с увеличением титра CYDV-RPV, но скорость передачи не зависит от поступления питательных веществ, стехиометрии тканей, структурных особенностей растений и титра вируса. Наши результаты показывают более сложную взаимосвязь между питательными веществами в окружающей среде, динамикой вирусов, изменениями фенотипа растений и конкуренцией между патогенами внутри хозяина.
Поставки питательных веществ из окружающей среды и химический состав тканей растений не повлияли напрямую на титр вируса
Теория экологической стехиометрии предсказывает, что соотношение в питательных веществах окружающей среды или содержании питательных веществ в организме хозяина ограничит успешность инфекции и производство вирусных частиц, требующих азота и фосфора (Sterner and Elser, 2002), тогда как теория конкуренции за ресурсы предсказывает рост популяции, устойчивость и сосуществование видов на основе как по норме питательных веществ, так и по соотношению. Нормы и соотношения поступления питательных веществ, в частности углерода, азота и фосфора, могут ограничивать размножение вирусов в водорослях и хозяевах фитопланктона (Elser et al., 2003; Класен и Эльзер, 2007; Frost et al., 2008; Маат и Брюссаард, 2016). В отличие от этих прогнозов и тестов в водных экосистемах, мы не обнаружили какого-либо прямого воздействия запасов питательных веществ или стехиометрии растительной ткани на плотность (т. Е. Титр) двух видов вирусов, изученных в нашем исследовании. Наше исследование показывает, что изменения фенотипических признаков растений могут быть ключевой связью между уровнем и соотношением питательных веществ и эпидемиологическими показателями.
Инфекция и источники питательных веществ из окружающей среды Измененный фенотип растения
В нашем исследовании BYDV-PAV и CYDV-RPV не снижали наземную биомассу, что контрастирует с другими исследованиями инфекции B / CYDV (Catherall, 1966; Malmstrom et al., 2005; Mordecai et al., 2015). Однако мы обнаружили, что масса листьев на площадь (то есть LMA) была выше у растений, инфицированных CYDV-RPV, что титр CYDV-RPV был выше у растений с высокими листьями LMA и что хлоротические симптомы, индуцированные на листьях, усиливались с CYDV-RPV. титр. Предыдущая работа над этой группой вирусов обнаружила снижение свежей биомассы и урожайности травяных хозяев, инфицированных B / CYDV, а инфицированные хозяева также характеризовались более высокой сухой массой листьев, что аналогично нашим результатам для растений, инфицированных CYDV-RPV (Jensen, 1968, 1969, 1972).Влияние B / CYDV на фенотип растений возникает из-за снижения транслокации фотосинтатов от листьев к корням и апикальной меристеме, что приводит к накоплению растворимых углеводов, крахмала и нерастворимых белков в пораженных листьях, увеличению сухой массы и скорости дыхания в темноте, снижению фотосинтеза. и индукция хлоротических симптомов (Jensen, 1968, 1969, 1972; Malmstrom and Field, 1997). Повышенная концентрация нерастворимого белка в инфицированных листьях может быть связана с накоплением нерастворимых белков, свободных аминокислот, нуклеиновых кислот, аминов, амидов и неорганического азота (Jensen, 1969).Эти результаты в сочетании с нашим исследованием предполагают, что наблюдаемый здесь более высокий LMA, связанный с инфекцией CYDV-RPV, может быть результатом снижения транслокации питательных веществ и последующего накопления C-богатых (например, углеводов и крахмала) и N-богатых (например, нерастворимых белковая фракция) соединений в инфицированных листьях. Накопление этих ресурсов в листьях инфицированных растений может вызвать более сильные изменения фенотипов, чем запасы питательных веществ, и может облегчить цикл размножения CYDV-RPV, повысить титр вируса и привести к снижению продукции фотосинтата и его транслокации в другие части растения, вызывая симптомы хлоротика.
Возможная роль соединений, богатых углеродом и азотом, в тканях растений в титре вируса
Наша работа также предполагает, что репликация CYDV-RPV может быть напрямую ограничена не общим содержанием и соотношениями азота, фосфора и углерода в тканях, а концентрацией и соотношением определенных молекул, богатых углеродом и / или азотом. Хотя инфекция B / CYDV нарушает транслокацию питательных веществ и увеличивает содержание соединений, богатых C и N в инфицированных листьях, большее количество углеводов накапливается в течение дня в листьях инфицированных по сравнению сБыло показано, что неинфицированные растения, выращенные в условиях повышенного содержания CO 2 , конвертируются, экспортируются и / или дышат в течение ночи из-за снижения концентрации сахарозы, глюкозы и фруктозы (Malmstrom and Field, 1997). Это говорит о том, что сахара могли быть перераспределены, чтобы способствовать размножению вирусов. Кроме того, было обнаружено, что накопление BYDV-PAV внутри хозяина увеличивалось у растений-хозяев, подвергшихся воздействию повышенных уровней CO 2 , хотя это увеличение не было связано ни с усилением роста растений, ни с абсолютной сухой массой (г), ни с средней площадью листьев (см ). -2 ) (Trebicki et al., 2015). Однако удельная площадь листа (см 2 г -1 ) была ниже (т. Е. Выше LMA) у инфицированных растений по сравнению с неинфицированными растениями (Trebicki et al., 2015), что указывает на потенциальную положительную корреляцию между титром BYDV-PAV и LMA. Напротив, усиление роста хозяина в условиях повышенного поступления азота привело к увеличению титра BYDV-PAV в другом исследовании (Whitaker et al., 2015), хотя это наблюдалось только у небольших растений, а у более крупных растений этот эффект был обратным (Whitaker et al. al., 2015), возможно, из-за уменьшения распределения питательных веществ растениями по клеточному механизму с увеличением размера растений (Elser et al., 2010). Наконец, углеводы (например, фруктоза, маннит и трегалоза) были выше как у здоровых, так и у инфицированных B / CYDV растений при повышенных уровнях CO 2 , а концентрации аминокислот были выше у инфицированных растений независимо от уровней CO 2 в атмосфере ( Vassiliadis et al., 2016). Взятые вместе, эти исследования показывают, что титр B / CYDV может больше зависеть от содержания и соотношений конкретных богатых C (например, CYDV).например, сахаров, таких как сахароза и фруктоза) и молекул, богатых азотом (например, аминокислот), чем концентрация элементарных питательных веществ как таковая . Таким образом, совместный анализ метаболических профилей у инфицированных и здоровых растений, накопления вируса внутри хозяина и передачи вируса между хозяином, а также молекулярных механизмов, лежащих в основе этих процессов, вероятно, представляет собой плодотворное направление для исследований.
Различия между вирусами и видами растений по функциональным признакам могут влиять на инфекцию и реакцию растений на поступление питательных веществ из окружающей среды
Мы не обнаружили никакой связи между титром или скоростью передачи BYDV-PAV и питательными веществами в окружающей среде, характеристиками растений или стехиометрией.В наших экспериментальных условиях нормы и соотношения питательных веществ в окружающей среде и тканях хозяина, возможно, не были ограничивающими для цикла размножения BYDV-PAV. Как предполагалось ранее, вопрос о потенциальных различиях в потребностях в питательных веществах для вирусов CYDV-RPV и BYDV-PAV остается открытым, и ограничения для CYDV-RPV и N-богатых метаболитов могут быть сильнее для CYDV-RPV, чем для BYDV-PAV ( Lacroix et al., 2014; Смит, 2014). Более того, Cronin et al. (2010) обнаружили межвидовые различия в способности видов растений действовать в качестве резервуаров BYDV-PAV, так что восприимчивость хозяина к инокуляции, накопление внутри хозяина и скорость передачи BYDV-PAV были в среднем выше для видов растений, характеризующихся «более быстрый» фенотип растения (Cronin et al., 2010). Кроме того, в то время как предыдущие полевые эксперименты продемонстрировали положительный эффект предложения P и соотношения N: P на распространенность B / CYDV (Borer et al., 2010, 2014b), повышенное снабжение P травяными хозяевами в контролируемых условиях уменьшало BYDV-PAV в пределах — накопление хозяев у Avena fatua , но не у травяных хозяев Bromus hordeaceus (Poaceae) (Rua et al., 2013). В нашем исследовании мы обнаружили различия во влиянии одиночного вируса CYDV-RPV, одиночного вируса BYDV-PAV и коинфекции на фенотип растений, что согласуется с формирующейся перспективой разнообразия возможных взаимодействий вируса-хозяина (Marquez et al., 2007; Roossinck et al., 2010; Русинк, 2015). Хотя один BYDV-PAV не оказывал значительного изменения фенотипа растений, коинфицированные растения характеризовались более высоким LMA, пониженным соотношением надземной и подземной биомассы и ростом растений, более высоким содержанием N и P; более высокое соотношение N: P; и больше стареющей ткани, чем неинфицированные или однократно инфицированные CYDV-RPV хозяева, что позволяет предположить, что коинфекция снижает способность растений выделять ресурсы для роста растений. В целом, эти результаты предполагают, что реакция растений на инфекцию, а также эпидемиологические параметры вируса на различных стадиях инфекции могут зависеть от множества факторов, включая меж- и внутривидовые различия в функциональных характеристиках вируса и растений, стехиометрию питательных веществ и метаболитов растений и абиотические условия окружающей среды.
Поставки питательных веществ из окружающей среды могут по-разному влиять на различные этапы эпидемиологического цикла
Было показано, что в наземных системах повышенное снабжение питательными веществами из окружающей среды видов трав в естественных экосистемах увеличивает распространенность и уровни коинфекции группы универсальных и переносимых тлей вирусов растений (т. Е. Вирусов желтых карликов ячменя и злаковых; Seabloom et al. др., 2009, 2013; Борер и др., 2010, 2014а, б). Эти реакции, наблюдаемые в естественных условиях, могут быть результатом множества взаимодействующих процессов, таких как изменения в сообществе растений-хозяев (Borer et al., 2009, 2010), измененное поведение или производительность переносчиков (Borer et al., 2009; Seabloom et al., 2013), прямое влияние снабжения растений питательными веществами и стехиометрии растений на восприимчивость хозяина, межвидовые взаимодействия вирусов, размножение вирусов и передачи (Смит, 1993, 2007; Смит и Холт, 1996), или через косвенное влияние на разнообразие видов, состав и функциональные характеристики растений (Liu et al., 2017).
Наши работы и работы, представленные Lacroix et al. (2014) теперь дают более глубокое представление о механизмах на уровне хозяина, которые лежат в основе воздействия питательных веществ на распространение патогенов, наблюдаемого в естественных экосистемах.Важно отметить, что мы показали, что на успех инфекции по-разному влияли скорость поступления питательных веществ и конкуренция вирусов (Lacroix et al., 2014). Уровень инфицирования CYDV-RPV был снижен как за счет объемов поставки P, так и за счет конкуренции с BYDV-PAV, но только при низких уровнях поступления азота. Это исследование показывает, что после установления инфекции титр CYDV-RPV был затронут из-за изменений фенотипа растений, возможно, из-за изменений скоростей и соотношений C-богатых и N-богатых метаболитов, а титр снижался из-за присутствия конкурента (коинфекция ).Более высокий титр вируса приводил к усилению проявления хлоротических симптомов, по крайней мере, для CYDV-RPV. В наших условиях передача, заключительный этап эпидемиологического цикла, не зависела от наличия питательных веществ, фенотипа хозяина, коинфекции вируса и титра вируса. Эксперимент по передаче в нашем исследовании был разработан для оценки влияния поступления питательных веществ из окружающей среды на передачу от хозяина к хозяину через изменение титра вируса. Титр вируса во всех хозяевах и / или период доступа к получению, позволяющий тлю приобретать вирусные частицы из ткани хозяина, мог быть достаточно высоким, чтобы максимизировать скорость передачи между хозяевами в наших экспериментальных условиях, что могло бы объяснить отсутствие корреляции между титром и передача часто наблюдается в системах растение-вирус (Froissart et al., 2010). Помимо титра вируса, на передачу вируса от хозяина к хозяину может сильно влиять поведение переносчика тлей, особенно для устойчиво передаваемых вирусов, которые требуют длительного времени для захвата тлей (Froissart et al., 2010; Ingwell et al., 2012; Smith , 2014; Blanc, Michalakis, 2016). Дальнейшие исследования, посвященные изучению того, могут ли источники питательных веществ в окружающей среде влиять на передачу вируса-хозяина через изменение привлекательности хозяина для насекомых-переносчиков и пищевого поведения тли, могут углубить наши знания о множественных путях, посредством которых ресурсы питательных веществ хозяина могут влиять на динамику вирусов растений.
Общие выводы
Наша работа подчеркивает проблемы понимания последствий повышенного отложения питательных веществ и измененных глобальных биогеохимических циклов (Tilman et al., 2001; Rockström et al., 2009) на экологию и эпидемиологию болезней. В то время как экологическая стехиометрия и теория ресурсной конкуренции могут служить отправной точкой для понимания влияния измененных норм и соотношений подачи питательных веществ на болезнь и могут эффективно прогнозировать некоторые звенья в цепи передачи, наша работа подчеркивает важность разделения роли конкретных C -богатые и богатые азотом метаболиты на титр растительных вирусов, а не сосредоточение внимания исключительно на общем поступлении, содержании или соотношении элементарных азота, фосфора и углерода.Кроме того, эта работа демонстрирует, что критически важно понимать молекулярные механизмы, которые приводят к фенотипическим изменениям хозяина, которые лежат в основе взаимодействий хозяин-вирус. Обладая более глубоким пониманием прямых и косвенных путей, с помощью которых скорость и соотношение поступления питательных веществ влияют на динамику патогенов, мы продолжаем строить прогнозное понимание влияния изменения условий окружающей среды на размножение и передачу вируса.
Авторские взносы
CL, ES и EB разработали этот исследовательский проект.CL проводил эксперименты с помощью других членов лаборатории. CL проанализировал данные и подготовил документ, а ES и EB внесли существенный вклад в статистический анализ и исправления рукописей. Все авторы одобрили эту версию рукописи и согласились нести ответственность за все аспекты этой работы.
Финансирование
Мы получили поддержку от программы NSF по экологии и эволюции инфекционных заболеваний (гранты DEB-1015805 и EF-12-41895) для EB и ES.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим Мисси Рудин, Курру Реннер, Абдулрахмана Гамама, Эми Кендиг, Аарона Дэвида и Эрика Линда, а также студентов и аспирантов за помощь в лаборатории. Мы также благодарим Марти Деккерса (Университет Северной Каролины) и Бенхэма Э. Л. Локхарта (Университет Миннесоты) за обмен опытом и советы по методам обнаружения и передачи вирусов. Мы благодарны Стюарту Грею и Дон М. Смит (Корнельский университет), а также Дмитрию Моллову (Клиника болезней растений, Университет Миннесоты) за обмен и содействие в получении изолятов вирусов BYDV-PAV и CYDV-RPV.Мы также благодарны Джорджиане Мэй, Роберту В. Стернеру и Джеймсу Б. Котнеру за предоставленный доступ к оборудованию для кПЦР, анализатора CHN и спектрофотометра Varian. Мы благодарим Эми Кендиг, Бенуа Моури и рецензентов за чтение и полезные комментарии к этой рукописи.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02116/full#supplementary-material
Список литературы
Аалто, С.Л., Декастекер, Э., Пулккинен, К. (2015). Трехсторонняя перспектива стехиометрических изменений взаимодействий паразит-хозяин. Trends Parasitol . 31, 333–340. DOI: 10.1016 / j.pt.2015.04.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Адам А., Габоржани Р., Тобиас И. и Кирали З. (1987). Влияние азотного питания на концентрацию вирусов, фосфолипидов и галактолипидов в листьях ячменя, инфицированных вирусом полосовой мозаики ячменя (bsmv). Ann. Прил. Биол . 110, 313–319. DOI: 10.1111 / j.1744-7348.1987.tb03261.x
CrossRef Полный текст | Google Scholar
Алквист П., Нуири А. О., Ли В. М., Кушнер Д. Б. и Дай Б. Т. (2003). Факторы-хозяева в репликации генома РНК-вируса с положительной цепью. Дж. Вирол . 77, 8181–8186. DOI: 10.1128 / JVI.77.15.8181-8186.2003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Александр, Х. М. (2010). Болезни в естественных популяциях растений, сообществах и экосистемах: понимание экологических и эволюционных процессов. Plant Dis. 94, 492–503. DOI: 10.1094 / PDIS-94-5-0492
CrossRef Полный текст | Google Scholar
Ализон, С., Херфорд, А., Мидео, Н., и Ван Баален, М. (2009). Эволюция вирулентности и гипотеза компромисса: история, текущее положение дел и будущее. J. Evol. Биол. 22, 245–259. DOI: 10.1111 / j.1420-9101.2008.01658.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Аль-Наими, Ф. А., Гаррет, К. А., и Бокус, В.W. (2005). Конкуренция, содействие и дифференциация ниш у двух листовых патогенов. Oecologia 143, 449–457. DOI: 10.1007 / s00442-004-1814-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Bawden, F. C., and Kassanis, B. (1950a). Некоторые эффекты питания хозяина на восприимчивость растений к заражению определенными вирусами. Ann. Прил. Биол. 37, 46–57. DOI: 10.1111 / j.1744-7348.1950.tb00949.x
CrossRef Полный текст | Google Scholar
Bawden, F.К. и Кассанис Б. (1950b). Некоторые эффекты питания растений-хозяев на размножение вирусов. Ann. Прил. Биол. 37, 215–228. DOI: 10.1111 / j.1744-7348.1950.tb01040.x
CrossRef Полный текст | Google Scholar
Бедхом, С., Агнью, П., Сидобре, К., и Михалакис, Ю. (2004). Нормы реакции вирулентности через пищевой градиент. Proc. R. Soc. B-Biol. Sci. 271, 739–744. DOI: 10.1098 / rspb.2003.2657
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Blanc, S., и Михалакис, Ю. (2016). Манипулирование хозяевами и переносчиками вирусами растений и воздействие окружающей среды. Curr. Opin. Insect Sci. 16, 36–43. DOI: 10.1016 / j.cois.2016.05.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Борер, Э. Т., Адамс, В. Т., Энглер, Г. А., Адамс, А. Л., Шуман, К. Б., и Сиблум, Э. У. (2009). Плодовитость тлей и нашествие пастбищ: ключ к успеху — история жизни захватчиков. Ecol. Прил. 19, 1187–1196. DOI: 10.1890 / 08-1205.1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бурер Э. Т., Хоссейни П. Р., Сиблум Э. У. и Добсон А. П. (2007). Вызванное патогеном изменение доминирования аборигенов в сообществе пастбищ. Proc. Natl. Акад. Sci. США . 104, 5473–5478. DOI: 10.1073 / pnas.0608573104
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Борер, Э. Т., Лейн, А. Л., и Сиблум, Э. У. (2016). Многомасштабный подход к болезням растений с использованием концепции метасообщества. Annu. Rev. Phytopathol. 54, 397–418. DOI: 10.1146 / annurev-phyto-080615-095959
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Борер, Э. Т., Сиблум, Э. У., Грюнер, Д. С., Харпол, У. С., Хиллебранд, Х., Линд, Э. М. и др. (2014a). Травоядные животные и питательные вещества контролируют разнообразие пастбищных растений за счет ограничения света. Природа 508, 517–520. DOI: 10.1038 / природа13144
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бурильщик, Э.Т., Сиблум, Э. У., Митчелл, К. Э. и Кронин, Дж. П. (2014b). Множественные питательные вещества и травоядные животные взаимодействуют, чтобы управлять разнообразием, продуктивностью, составом и инфекцией на сукцессионных пастбищах. Oikos 123, 214–224. DOI: 10.1111 / j.1600-0706.2013.00680.x
CrossRef Полный текст | Google Scholar
Борер Э. Т., Сиблум Э. У., Митчелл К. Э. и Пауэр А. Г. (2010). Местный контекст способствует заражению трав универсальными трансмиссивными вирусами. Ecol.Lett. 13, 810–818. DOI: 10.1111 / j.1461-0248.2010.01475.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кардинале, Б. Дж., Хиллебранд, Х., Харпол, В. С., Гросс, К., и Птачник, Р. (2009). Отделение влияния «доступности» ресурсов от «дисбаланса» ресурсов на взаимосвязь между производительностью и разнообразием. Ecol. Lett. 12, 475–487. DOI: 10.1111 / j.1461-0248.2009.01317.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Катералл, П.Л. (1966). Влияние вируса желтой карликовости ячменя на рост и урожайность одиночных растений и искусственных травянистых растений ржи многолетней. Annu. Прил. Биол. 57, 155–162. DOI: 10.1111 / j.1744-7348.1966.tb06876.x
CrossRef Полный текст | Google Scholar
Чейн, Ф., Рио, Г., Тротте, М., и Жако, Э. (2005). Анализ закономерностей накопления вируса желтой карликовости ячменя-pav (bydv-pav) в двух устойчивых линиях пшеницы. Eur. J. Plant Pathol. 113, 343–355.DOI: 10.1007 / s10658-005-7966-7
CrossRef Полный текст | Google Scholar
Класен, Дж. Л., и Эльзер, Дж. Дж. (2007). Влияние соотношения углерод: фосфор хозяина chlorella nc64a на продукцию вируса-1 парамециума бурсария chlorella virus-1. Freshw. Биол. 52, 112–122. DOI: 10.1111 / j.1365-2427.2006.01677.x
CrossRef Полный текст | Google Scholar
Corner, E. D. S., Head, R. N., Kilvingston, C. C., and Pennycuick, L. (1976). О питании и обмене веществ зоопланктона. J. Mar. Biol. Доц. 56, 345–358. DOI: 10.1017 / S0025315400018968
CrossRef Полный текст | Google Scholar
Крейн, Дж. М., Тилман, Д., Ведин, Д., Райх, П., Тьелкер, М., и Кнопс, Дж. (2002). Функциональные особенности, продуктивность и влияние на круговорот азота 33 видов пастбищ. Funct. Ecol. 16, 563–574. DOI: 10.1046 / j.1365-2435.2002.00660.x
CrossRef Полный текст | Google Scholar
Кронин, Дж. П., Уэлш, М. Э., Деккерс, М.Г., Аберкромби, С. Т., и Митчелл, К. Э. (2010). Физиологический фенотип хозяина объясняет резервуарный потенциал патогенов. Ecol. Lett. 13, 1221–1232. DOI: 10.1111 / j.1461-0248.2010.01513.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дарси, К. Дж., И Бернетт, П. А. (1995). Ячменный желтый карлик, 40 лет прогресса . Сент-Пол, Миннесота: Американское фитопатологическое общество.
Деб М. и Андерсон Дж. М. (2008). Разработка метода множественного обнаружения ПЦР для вирусов желтой карликовости ячменя и злаковых, вируса полосатости пшеницы, вируса мозаики полосатой пшеницы и вируса мозаики пшеницы, передаваемого через почву. J. Virol. Методы 148, 17–24. DOI: 10.1016 / j.jviromet.2007.10.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дордас, К. (2009). «Роль питательных веществ в борьбе с болезнями растений в устойчивом сельском хозяйстве: обзор», в Sustainable Agriculture , ред. Э. Лихтфуз, М. Наваррете, П. Дебаеке, С. Вероник и К. Альберола (Дордрехт: Springer), 443 –460.
Google Scholar
Doumayrou, J., Leblaye, S., Froissart, R., и Михалакис Ю. (2013). Уменьшение площади листьев и тяжести симптомов как индикаторы смертности растений, вызванной заболеванием: пример вируса мозаики цветной капусты, заражающего двух хозяев Brassicaceae. Virus Res. 176, 91–100. DOI: 10.1016 / j.virusres.2013.05.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Даунс, Р. Дж., И Хеллмерс, Х. (1975). Окружающая среда и экспериментальный контроль роста растений . Нью-Йорк, штат Нью-Йорк: Academic Press.
Google Scholar
Эльзер, Дж.J., Acharya, K., Kyle, M., Cotner, J., Makino, W., Markow, T., et al. (2003). Связь между скоростью роста и стехиометрией в разнообразной биоте. Ecol. Lett. 6, 936–943. DOI: 10.1046 / j.1461-0248.2003.00518.x
CrossRef Полный текст | Google Scholar
Эльзер, Дж. Дж., Бракен, М. Е. С., Клеланд, Е. Е., Грюнер, Д. С., Харпол, В. С., Хиллебранд, Х. и др. (2007). Глобальный анализ ограничения азота и фосфора первичными продуцентами в пресноводных, морских и наземных экосистемах. Ecol. Lett. 10, 1135–1142. DOI: 10.1111 / j.1461-0248.2007.01113.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эльзер, Дж. Дж., Фаган, В. Ф., Керкхофф, А. Дж., Свенсон, Н. Г., и Энквист, Б. Дж. (2010). Биологическая стехиометрия производства растений: метаболизм, масштабирование и экологическая реакция на глобальные изменения. New Phytol. 186, 593–608. DOI: 10.1111 / j.1469-8137.2010.03214.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ераслан, Ф., Акбас, Б., Инал, А., и Таракчиоглу, К. (2007). Влияние заражения вирусом томатной мозаики (tomv) источниками кальция с опрыскиванием на листьях растений томата, выращиваемых в теплицах. Phytoparasitica 35, 150–158. DOI: 10.1007 / BF02981110
CrossRef Полный текст | Google Scholar
Фруассар Р., Думайру Дж., Вийом Ф., Алисон С. и Михалакис Ю. (2010). Компромисс между вирулентностью и передачей трансмиссивных вирусов растений: обзор (не) существующих исследований. Philos.Пер. R. Soc. B Biol. Sci. 365, 1907–1918. DOI: 10.1098 / rstb.2010.0068
PubMed Аннотация | CrossRef Полный текст
Фрост П. К., Эберт Д. и Смит В. Х. (2008). Ответы бактериального патогена на ограничение фосфора его водным беспозвоночным хозяином. Экология 89, 313–318. DOI: 10.1890 / 07-0389.1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Грюбер К. Э., Накагава С., Лоус Р. Дж. И Джеймисон И. Г. (2011).Многомодельный вывод в экологии и эволюции: проблемы и решения. J. Evol. Биол. 24, 699–711. DOI: 10.1111 / j.1420-9101.2010.02210.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Halbert, S., and Voegtlin, D. (1995). «Биология и таксономия переносчиков вирусов желтых карликов ячменя», в Barley Yellow Dwarf, 40 Years of Progress , ред. К. Дж. Дарси и П. А. Бернетт (Сент-Пол, Миннесота: Американское фитопатологическое общество), 217–258.
Гессен, Д. О., Эльзер, Дж. Дж., Стернер, Р. В., и Урабе, Дж. (2013). Экологическая стехиометрия: элементарный подход с использованием основных принципов. Лимнол. Oceanogr. 58, 2219–2236. DOI: 10.4319 / lo.2013.58.6.2219
CrossRef Полный текст | Google Scholar
Хиллебранд, Х., Коулз, Дж. М., Левандовска, А., Ван де Ваал, Д. Б., и Слива, К. (2014). Подумайте о соотношении! Стехиометрический взгляд на исследования функционирования экосистемы биоразнообразия. Basic Appl. Ecol. 15, 465–474.DOI: 10.1016 / j.baae.2014.06.003
CrossRef Полный текст | Google Scholar
Хоугланд Д. Р. и Арнон. Д. И. (1938). Метод водной культуры для выращивания растений без почвы. Калифорнийская экспериментальная сельскохозяйственная станция, циркуляр 347, 1:32.
Google Scholar
Ирвин М. Э. и Треш Дж. М. (1990). Эпидемиология желтого карлика ячменя — исследование в экологической сложности. Annu. Rev. Phytopathol. 28, 393–424. DOI: 10.1146 / annurev.py.28.0
.002141
CrossRef Полный текст | Google Scholar
Дженсен, С. Г. (1968). Фотосинтез, дыхание и другие физиологические взаимоотношения у карликового вируса ячменя. Фитопатология 58, 204–208.
Дженсен, С. Г. (1969). Состав и метаболизм листьев ячменя, инфицированных вирусом желтой карликовости ячменя. Фитопатология 59, 1694–1698.
Дженсен, С. Г. (1972). Метаболизм и углеводный состав пшеницы, инфицированной вирусом желтой карликовости ячменя. Фитопатология 62, 587–592. DOI: 10.1094 / Phyto-62-587
CrossRef Полный текст
Джовер, Л. Ф., Эффлер, Т. К., Бьюкен, А., Вильгельм, С. В., и Вайц, Дж. С. (2014). Элементный состав вирусных частиц: последствия для морских биогеохимических циклов. Nat. Rev. Microbiol. 12, 519–528. DOI: 10.1038 / nrmicro3289
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Карпинец, Т. В., Гринвуд, Д. Дж., Сэмс, К. Э., и Аммонс, Дж.Т. (2006). Соотношение РНК: белок в одноклеточном организме как характеристика стехиометрии фосфора и азота и потребности клетки в рибосомах для синтеза белка. BMC Biol. 4:30. DOI: 10.1186 / 1741-7007-4-30
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лакруа К., Сиблум Э. У. и Борер Э. Т. (2014). Обеспечение питательными веществами окружающей среды изменяет распространенность и ослабляет конкурентное взаимодействие между коинфекционными вирусами. New Phytol. 204, 424–433. DOI: 10.1111 / nph.12909
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ланге, Б., Рейтер, М., Эберт, Д., Муйларт, К., и Декастекер, Э. (2014). Качество диеты определяет межвидовые взаимодействия паразитов в популяциях хозяев. Ecol. Evol. 4, 3093–3102. DOI: 10.1002 / ece3.1167
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Leclercq-Le Quillec, F., Plantegenest, M., Riault, G., and Dedryver, C.А. (2000). Анализ и моделирование темпорального развития серотипов вируса желтой карликовости ячменя на ячменных полях. Фитопатология 90, 860–866. DOI: 10.1094 / PHYTO.2000.90.8.860
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лю, X., Лю, С., Сан, Д., Брэдшоу, К. Дж. А., и Чжоу, С. (2017). Сокращение численности видов при азотных удобрениях повышает уровень осведомленности о грибковых заболеваниях на уровне сообществ. Proc. R. Soc. B Biol. Sci. 284: 2016 2621.DOI: 10.1098 / rspb.2016.2621
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маат, Д. С., Брюссар, К. П. Д. (2016). Ограничение как фосфора, так и азота сдерживает распространение вирусов в морском фитопланктоне. Aquat. Microb. Ecol. 77, 87–97. DOI: 10.3354 / ame01791
CrossRef Полный текст | Google Scholar
Макартур Р. Х. (1972). Географическая экология: закономерности в распределении видов . Принстон, Нью-Джерси: Издательство Принстонского университета.
Google Scholar
Мальмстрем, К. М., и Филд, К. Б. (1997). Различия в реакции растений овса на повышенный уровень углекислого газа, вызванные вирусами. Plant Cell Environ. 20, 178–188. DOI: 10.1046 / j.1365-3040.1997.d01-63.x
CrossRef Полный текст | Google Scholar
Мальмстрем, К. М., Хьюз, К. К., Ньютон, Л. А., и Стоунер, К. Дж. (2005). Вирусная инфекция в остатках местных кустарниковых трав с захваченных лугов Калифорнии. New Phytol. 168, 217–230.DOI: 10.1111 / j.1469-8137.2005.01479.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маркес, Л. М., Редман, Р. С., Родригес, Р. Дж., И Русинк, М. Дж. (2007). Вирус в грибке на растении: трехсторонний симбиоз, необходимый для термостойкости. Science 315, 513–515. DOI: 10.1126 / science.1136237
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мауле А., Лех В. и Ледерер К. (2002). Диалог между вирусами и хостами в совместимых взаимодействиях. Curr. Opin. Plant Biol. 5, 279–284. DOI: 10.1016 / S1369-5266 (02) 00272-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Миллер Т. Э., Бернс Джин Х., Мунгиа П., Уолтерс Эрик Л., Кнейтель Джейми М., Ричардс Пол М. и др. (2005). Критический обзор двадцатилетнего использования теории соотношения ресурсов. Am. Nat. 165, 439–448. DOI: 10.1086 / 428681
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Митчелл, К.Э., Райх П. Б., Тилман Д. и Грот Дж. В. (2003). Влияние повышенного содержания CO2, осаждения азота и уменьшения видового разнообразия на листовые грибковые заболевания растений. Glob. Сменить Биол. 9, 438–451. DOI: 10.1046 / j.1365-2486.2003.00602.x
CrossRef Полный текст | Google Scholar
Мордехай, Э. А., Хинденланг, М., Митчелл, К. Э. (2015). Различное влияние разнообразия вирусов на продукцию биомассы местной и экзотической травы-хозяина. PLoS ONE 10: e0134355.DOI: 10.1371 / journal.pone.0134355
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перри К. Л., Колб Ф. Л., Сэммонс Б., Лоусон К., Цисар Г. и Ом Х. (2000). Влияние вируса желтой карликовости ячменя на урожайность мягкой красной озимой пшеницы. Фитопатология 90, 1043–1048. DOI: 10.1094 / PHYTO.2000.90.9.1043
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пауэр А. Г. и Грей С. М. (1995). «Передача тлей вирусов желтых карликов ячменя: взаимодействие между вирусами, переносчиками и растениями-хозяевами», в «Желтый карлик ячменя: 40 лет прогресса» , ред.Дж. Дарси и П. А. Бернетт (Сент-Пол, Миннесота: Американское фитопатологическое общество), 259–289.
Редфилд, А. К. (1958). Биологический контроль химических факторов в окружающей среде. Am. Sci. 46, 205–221.
Google Scholar
Райх, П. Б. (2014). Мировой спектр экономики растений «быстро-медленно»: манифест черт. J. Ecol. 102, 275–301. DOI: 10.1111 / 1365-2745.12211
CrossRef Полный текст | Google Scholar
Риго, Т., Перро-Миннот, М. Дж., И Браун, М. Дж. Ф. (2010). Группы паразитов и хозяев: принятие реальности улучшит наши знания о передаче паразитов и вирулентности. Proc. R. Soc. B Biol. Sci. 277, 3693–3702. DOI: 10.1098 / rspb.2010.1163
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Робертсон, Н. Л., и Френч, Р. (2007). Генетическая структура природных популяций изолятов вируса желтой карликовости ячменя / злаков с Аляски. Arch. Virol. 152, 891–902.DOI: 10.1007 / s00705-006-0913-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Rockström, J., Steffen, W., Noone, K., Persson, A., Chapin, F. S., Lambin, E. F., et al. (2009). Безопасное рабочее пространство для человечества. Nature 461, 472–475. DOI: 10.1038 / 461472a
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Roossinck, M.J., Saha, P., Wiley, G.B., Quan, J., White, J.D., Lai, H., et al. (2010). Экогеномика: использование массового параллельного пиросеквенирования для понимания экологии вирусов. Mol. Ecol. 19, 81–88. DOI: 10.1111 / j.1365-294X.2009.04470.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Руа, М. А., Умбанховар, Дж., Ху, С., Берки, К. О., Митчелл, К. Э. (2013). Повышенный уровень CO2 стимулирует взаимные положительные эффекты между растительным вирусом и арбускулярным микоризным грибком. New Phytol. 199, 541–549. DOI: 10.1111 / nph.12273
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Salvaudon, L., Де Мораес, К. М., и Мешер, М. К. (2013). Результаты совместного заражения двумя потивирусами: последствия для эволюции манипулятивных стратегий. Proc. R. Soc. B Biol. Sci. 280: 20122959. DOI: 10.1098 / rspb.2012.2959
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сиблум, Э. У., Борер, Э. Т., Бакли, Ю. М., Клеланд, Э. Э., Дэвис, К. Ф., Фирн, Дж. И др. (2015a). Происхождение видов растений предсказывает доминирование и реакцию на обогащение питательными веществами и травоядных животных на глобальных пастбищах. Nat. Commun. 6, 7710. DOI: 10.1038 / ncomms8710
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сиблум, Э. У., Борер, Э. Т., Гросс, К., Кендиг, А. Э., Лакруа, К., Митчелл, К. Э. и др. (2015b). Экология сообщества патогенов: совместное заражение, сосуществование и состав сообщества. Ecol. Lett. 18, 401–415. DOI: 10.1111 / ele.12418
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сиблум, Э. У., Борер, Э.Т., Лакруа К., Митчелл К. Э. и Пауэр А. Г. (2013). Богатство и состав сообществ вирусных патогенов, собранных в нишах. PLoS ONE 8: e55675. DOI: 10.1371 / journal.pone.0055675
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сиблум, Э. У., Борер, Э. Т., Митчелл, К. Э., и Пауэр, А. Г. (2010). Вирусное разнообразие и градиенты распространенности на лугах тихоокеанского побережья Северной Америки. Экология 91, 721–732. DOI: 10.1890 / 08-2170.1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сиблум, Э.У., Хоссейни П. Р., Пауэр А. Г. и Борер Э. Т. (2009). Разнообразие и состав вирусных сообществ: совместное заражение вирусами желтых карликов ячменя и злаковых на лугах Калифорнии. Am. Nat. 173, E79 – E98. DOI: 10.1086 / 596529
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Смит В. Х., Джонс Т. П. и Смит М. С. (2005). Питание хозяина и инфекционные заболевания: экологический взгляд. Фронт. Ecol. Environ. 3, 268–274. DOI: 10.1890 / 1540-9295 (2005) 003 [0268: HNAIDA] 2.0.CO; 2
CrossRef Полный текст | Google Scholar
Стернер Р. У. и Эльзер Дж. Дж. (Ред.). (2002). Экологическая стехиометрия: биология элементов от молекул до биосферы . Принстон, Нью-Джерси: Издательство Принстонского университета.
Google Scholar
Тилман Д. (1976). Экологическая конкуренция между водорослями — экспериментальное подтверждение теории ресурсной конкуренции. Science 192, 463–465.DOI: 10.1126 / science.192.4238.463
CrossRef Полный текст | Google Scholar
Тилман Д. (1977). Ресурсная конкуренция между планктонными водорослями: экспериментальный и теоретический подход. Экология 58, 338–348. DOI: 10.2307 / 1935608
CrossRef Полный текст | Google Scholar
Тилман Д. (1982). Конкуренция ресурсов и структура сообщества . Принстон, Нью-Джерси: Издательство Принстонского университета.
Google Scholar
Тильман, Д., Фарджоне, Дж., Вольф, Б., Д’Антонио, К., Добсон, А., Ховарт, Р. и др. (2001). Прогнозирование глобальных изменений окружающей среды, вызванных сельским хозяйством. Science 292, 281–284. DOI: 10.1126 / science.1057544
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Требицки П., Нанкарроу Н., Коул Э., Боск-Перес Н. А., Констебль Ф. Э., Фриман А. Дж. И др. (2015). Согласно прогнозам, количество вирусов в пшенице будет увеличиваться с изменением климата. Glob. Сменить Биол. 21, 3511–3519.DOI: 10.1111 / gcb.12941
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тернер П. Э. (2005). Паразитизм между одновременно инфицированными бактериофагами. Adv. Ecol. Res. 37, 309–332. DOI: 10.1016 / S0065-2504 (04) 37010-8
CrossRef Полный текст | Google Scholar
Вассилиадис, С., Пламмер, К. М., Пауэлл, К. С., Требицки, П., Удача, Дж. Э. и Рочфорт, С. Дж. (2016). Влияние повышенного CO 2 и вирусной инфекции на первичный метаболизм пшеницы. Funct. Plant Biol. 43, 892–902. DOI: 10.1071 / FP15242
CrossRef Полный текст | Google Scholar
Виолле, К., Навас, М. Л., Вайл, Д., Казаку, Э., Фортунель, К., Хаммель, И. и др. (2007). Пусть понятие черты будет функциональным !. Oikos 116, 882–892. DOI: 10.1111 / j.0030-1299.2007.15559.x
CrossRef Полный текст | Google Scholar
Виолле, К., Райх, П. Б., Пакала, С. В., Энквист, Б. Дж., И Каттге, Дж. (2014). Возникновение и перспективы функциональной биогеографии. Proc. Natl. Акад. Sci. США 111, 13690–13696. DOI: 10.1073 / pnas.1415442111
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вестоби, М. (1998). Схема стратегии экологии растений с высотой листа и семенами (lhs). Растительная почва 199, 213–227. DOI: 10.1023 / A: 1004327224729
CrossRef Полный текст | Google Scholar
Уитакер, Б. К., Руа, М. А., и Митчелл, К. Э. (2015). Производство вирусных патогенов в дикорастущих травах-хозяевах обусловлено ростом хозяев и почвенным азотом. New Phytol. 207, 760–768. DOI: 10.1111 / nph.13369
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Райт, И. Дж., Райх, П. Б., Вестоби, М., Акерли, Д. Д., Барух, З., Бонгерс, Ф., и др. (2004). Спектр листовой экономики во всем мире. Nature 428, 821–827. DOI: 10.1038 / nature02403
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цендер, К. Б., и Хантер, М. Д. (2009). Больше не обязательно лучше: влияние ограниченных и чрезмерных питательных веществ на темпы роста популяции травоядных. Ecol. Энтомол. 34, 535–543. DOI: 10.1111 / j.1365-2311.2009.01101.x
CrossRef Полный текст | Google Scholar
Обзор питательных веществ
Витамины подразделяются на водорастворимые и жирорастворимые. В организме человека 15 витаминов: 5 жирорастворимых (A, D, E и K1, K2) и 10 водорастворимых (9 витаминов B и витамин C).
Ценность употребления определенной пищи для поддержания здоровья была признана задолго до того, как были идентифицированы эти витамины.
Известно, что в 1747 году шотландский хирург Джеймс Линд обнаружил, что продукты из цитрусовых излечивают цингу, особенно смертельное заболевание, при котором коллаген не образуется должным образом, вызывая плохое заживление ран, кровотечение десен, сильную боль и смерть.
В 1753 году Линд опубликовал свой «Трактат о цинге», в котором рекомендовал использовать лимоны и лайм не только для лечения, но и для предотвращения цинги, что значительно позже было принято британским королевским флотом.
Позже было обнаружено, что витамин С является жизненно важным микронутриентом, покончившим с цингой.
В конце 19 — начале 20 веков использование исследований депривации позволило ученым выделить и идентифицировать ряд витаминов.
Липид из рыбьего жира использовался для лечения рахита у крыс, а жирорастворимое питательное вещество было названо «антирахитическим средством А». Таким образом, первый когда-либо выделенный «витаминный» биоактивный препарат, излечивающий рахит, первоначально был назван «витамином А»; однако биоактивность этого соединения теперь называется витамином D.
Минералы в основном используются организмом для создания крепких костей и зубов, контроля жидкостей организма внутри и вне клеток, регулирования нашего сердцебиения и превращения пищи, которую мы едим, в энергию.
Минералы также поддерживают внутренний баланс нашего тела, поэтому они остаются в стабильном и здоровом состоянии при изменении внешних условий вокруг нас.
Минералы играют важную роль в коммуникации между нашими клетками, например, кальций действует как посредник между клетками нашего мозга.
Фитохимические вещества или полифенолы
Фитохимические вещества / полифенолы — это химические вещества, производимые растениями.Это химические вещества, которые растения используют для защиты от болезней.
Например, растения защищают себя от бактерий, грибков, вирусов и повреждения клеток фитостеринами, каротиноидами, флавоноидами, изотиоцианатами и подобными химическими веществами.
Исследования показали, что эти же химические вещества могут оказаться одинаково полезными для человеческого организма, помогая в таких областях, как рак, болезни сердца и профилактика диабета.
Национальный институт рака определил в лабораторных исследованиях, что более 1000 различных фитохимических веществ могут обладать профилактической активностью.
Подсчитано, что всего в одной порции овощей может содержаться более 100 различных фитохимических веществ. Для оценки способности определенных съедобных растений предотвращать рак были проведены многочисленные исследования на клеточных культурах и на животных моделях.
Еще в 1980 году программа химиопрофилактики NCI начала оценку фитохимических веществ на предмет безопасности, эффективности и применимости для профилактики рака.
Майкл Спорн ввел термин «химиопрофилактика» в середине 1970-х годов для описания стратегии блокирования или замедления возникновения опухолей с помощью относительно нетоксичных химических веществ.
NCI проводит исследования более 400 потенциальных агентов и спонсирует более 65 испытаний химиопрофилактики фазы I, фазы II и фазы III. Сюда входят различные вещества или их смеси, многие из которых являются фитохимическими веществами пищевого происхождения.
Фитохимические вещества часто считаются антиоксидантами, однако их антиоксидантная (очищающая) активность гораздо менее сильна, чем их положительное влияние на передачу сигналов в клетках. Передача сигналов здоровыми клетками жизненно важна для предотвращения искажений, ведущих к раку.
Фитохимические вещества на сайте checkyourfood.com сгруппированы по флавоноидам, изофлавонам, фитостеринам, ликопену, лютеину и зеаксантину.
Мы также выделили некоторые специфические фитохимические вещества из этих групп, такие как кверцетин, поскольку исследования проводились именно с ними.
Исследования неизменно показывают, что диета с высоким содержанием фруктов и овощей снижает риск хронических заболеваний, однако сложные взаимодействия между признанными и неидентифицированными фитохимическими веществами еще полностью не изучены.
Доказательства того, что очень высокие дозы в виде добавок отдельных фитохимических веществ являются полезными, противоречивы и являются слабым аргументом в пользу естественного питания, чтобы максимизировать еще не полностью изученные взаимодействия между фитохимическими веществами.
Жиры также являются богатым источником энергии, ключевым компонентом клеточных мембран, сигнальных молекул и изоляцией для наших нейронов (клетки, которая передает информацию по нашему телу).
Они также играют ключевую роль в транспортировке и хранении питательных веществ.Аминокислоты влияют на работу органов, желез, сухожилий и артерий.
Незаменимые аминокислоты жизненно важны для заживления ран и восстановления тканей, особенно в мышцах, костях, коже и волосах, а также для удаления всех видов отходов, образующихся в результате нашего метаболизма.
У нас есть набор доступных бесплатных аминокислот, которые наш организм может использовать в любое время. Этот пул меняется три-четыре раза в день, поэтому его необходимо поддерживать, чтобы наш метаболизм функционировал на пике.
Недавно ученый-исследователь из Института клеточной терапии и иммунологии им. Фраунгофера IZI в Лейпциге, Германия, обнаружил, что простые натуральные аминокислоты работают лучше, чем антибиотики, при лечении инфекций и не причиняют вреда здоровым клеткам в организме. Это вероятная причина эффективности желе из куриных лапок, которое мой отец давал при коклюше.
Химическое плодородие почв: питательные вещества почвы и питание растений
Скачать PDF
Кристофер Джонс
Менеджер по исследованиям
Программа исследований Северной АвстралииКлючевые моменты
- Есть ряд питательных элементов, которые необходимы для роста растений.
- Углерод, водород и кислород, три основных питательных вещества для растений, поступают из атмосферы и из воды. Другие важные питательные вещества поступают из почвы.
- Основными питательными веществами или макроэлементами, поступающими из почвы, являются азот, фосфор, калий, сера, магний и кальций.
- Незначительные питательные вещества, также называемые микроэлементами или микроэлементами, поставляемые почвой, включают молибден, медь, цинк, марганец, железо, никель, бор и хлор.
- Дефицит любого из этих важных питательных веществ замедлит рост, поскольку урожай растений зависит от количества наиболее ограниченного питательного вещества, каким бы оно ни было.
- Плодородие почвы представляет собой сложное взаимодействие между тремя компонентами плодородия: химическим плодородием, структурным плодородием и биологическим плодородием. На них сильно влияет присутствие углерода и воды.
- Наше понимание того, как взаимодействуют компоненты плодородия почвы, является неполным и требует дальнейших исследований, чтобы дать соответствующие рекомендации сельхозпроизводителям.
Сводка
Рост, сила роста и урожайность растений зависят от наличия ряда основных питательных веществ. Подробные списки этих элементов немного различаются, но большинство источников идентифицируют три основных неминеральных элемента: углерод, кислород и водород; шесть основных или макроэлементов: азот, фосфор, калий, сера, кальций и магний; и ряд микроэлементов, которые включают молибден, медь, бор, марганец, железо, никель, хлор и цинк.В дополнение к этим элементам есть также питательные вещества, которые либо необходимы некоторым, но не всем растениям, либо полезны, но не необходимы. Поглощение растениями всех питательных веществ ограничено единственным дефицитным элементом, а не наличием отдельного элемента.
Многие типы почв, встречающиеся в Австралии, испытывают дефицит некоторых основных питательных веществ, и решение не так просто, как просто добавить удобрения. На способность растения поглощать питательные вещества, даже если они имеются в изобилии, сильно влияет доступность воды, присутствие органических веществ в почве, структура почвы и микробиология.
Анализ
Введение
Более 150 лет назад наука определила, что некоторые химические элементы абсолютно необходимы для жизни растений. Сегодня, в зависимости от источника исследования, наука признает 16 или 17 элементов необходимыми для всех растений. Также были идентифицированы дополнительные элементы, которые либо необходимы для некоторых растений, либо полезны, но не важны для роста растений. Несмотря на незначительные различия во мнениях источников, это твердо установленный принцип, согласно которому почвы должны обеспечивать растения значительным количеством питательных веществ, содержащих химические элементы, чтобы растения могли завершить свой жизненный цикл.
В естественных, здоровых экосистемах уровни питательных веществ в почве поддерживаются круговоротом питательных веществ и являются относительно стабильными. Однако сельскохозяйственные почвы могут испытывать дефицит питательных веществ, поскольку сельскохозяйственные экосистемы не закрыты, а питательные вещества навсегда покидают систему в виде продуктов растительного или животного происхождения. Даже при использовании наилучших сельскохозяйственных методов такая открытая система не может существовать бесконечно. Доступ к питательным веществам для растений, их усвоение, замена и добавление не так просты, как определение и добавление химикатов.Механизмы поглощения разнообразны и могут быть сложными. Научные исследования продолжают улучшать наше понимание того, как наилучшим образом поддерживать химическое плодородие почвы.
Определение питательных веществ
Определение питательного вещества для растений обманчиво простое; это химический элемент, который необходим или полезен для роста растений или некоторых других процессов жизненного цикла, таких как размножение. Для того чтобы питательное вещество было классифицировано как Essential , оно должно быть либо необходимо для завершения жизненного цикла растения, либо должно быть частью жизненно важного компонента растения, такого как клеточные структуры или метаболический процесс, химические процессы, происходящие внутри живые организмы, например фотосинтез.Другими словами, для роста и размножения растения необходим существенный элемент (постулат Коха).
Основные питательные элементы
Из 17 основных питательных веществ углерода , кислорода и водорода классифицируются как неминеральные элементы. Доступ к ним осуществляется из воздуха и воды, поэтому они не считаются питательными веществами почвы. Углерод составляет основу многих биологических молекул растений, включая белки, крахмалы и целлюлозу.Он фиксируется фотосинтезом (процессом, при котором зеленые растения используют солнечный свет и хлорофилл для создания продуктов питания из углекислого газа и воды) из углекислого газа и является частью сахаров и крахмалов, которые хранят энергию в растении. Почти полностью водород получают из воды. Это важный элемент фотосинтеза и дыхания, процесса выработки энергии за счет потребления продуктов, полученных в результате фотосинтеза. Кислород получают из воздуха в виде газообразного кислорода или в молекулах воды или углекислого газа.Он также необходим для дыхания растений (процесса, при котором растение получает энергию от сахаров и крахмалов в присутствии кислорода). Остальные 14 элементов относятся к категории питательных веществ для почвы. Они делятся на две категории: макроэлементы и микроэлементы.
Макронутриенты, как следует из названия, необходимы в относительно больших количествах. Они необходимы для основных повседневных биологических функций растений, таких как рост, фотосинтез и дыхание.Макроэлементы:
- Азот необходим для всех процессов роста растений.
- Фосфор является важным компонентом многих жизненно важных производственных процессов.
- Калий также необходим для широкого круга важных процессов на предприятии.
- Сера требуется для образования нескольких аминокислот, белков и витаминов, а также для производства хлорофилла.
- Кальций участвует в правильном функционировании точек роста, особенно кончиков корней.
- Магний является важным компонентом хлорофилла и, следовательно, жизненно важен для фотосинтеза.
Микроэлементы, не менее важные, чем макронутриенты, необходимы только в небольших количествах. Дефицит питательных микроэлементов чаще встречается в сильно выщелоченных песках, органических почвах и в сильно щелочных почвах.Недостатки также могут развиваться на сильно возделываемой почве. Микроэлементы могут быть вредными или вредными для роста растений, если они присутствуют в больших количествах. Публикация CSIRO, Австралийские почвы и ландшафты; Иллюстрированный компендиум отмечает, что сельскохозяйственное развитие больших территорий Австралии было возможно только тогда, когда дефицит питательных микроэлементов был обнаружен и устранен. В настоящее время признанными основными микроэлементами являются:
- Молибден , который принимает непосредственное участие в метаболизме азота.
- Медь , необходимая для образования ферментов для производства хлорофилла.
- Бор необходим для движения сахаров по растению и метаболизма азота.
- Марганец, железо и цинк необходимы для процесса роста растений.
- Никель является самым недавно идентифицированным важным питательным веществом для растений. Это ключевой компонент процессов, участвующих в метаболизме азота и биологической фиксации азота.
- Хлор необходим для углеводного обмена и производства хлорофилла. Следует отметить, что хлор можно определить как макроэлемент. Из-за его обычного обилия в окружающей среде он очень редко бывает дефицитным и поэтому часто сгруппирован с питательными микроэлементами.
Помимо основных питательных веществ, есть ряд элементов, которые не полностью соответствуют строгому определению основных, но, тем не менее, важны.Эти элементы либо необходимы для некоторых, но не для всех видов растений, либо они очень полезны для роста растений. Важно отметить, что по мере роста наших научных знаний о питании растений растет и этот список элементов. Некоторые из питательных веществ в этой категории:
- Натрий , который в надлежащих количествах играет важную полезную роль в метаболизме растений. Это важно для небольшой группы растений, которые растут в среде с высоким содержанием соли, известных как галофиты. Он также полезен как частичный заменитель калия у некоторых видов.
- Кобальт оказался важным для эффективной фиксации атмосферного азота в корневых клубеньках бобовых (фасоль, чечевица и другие бобовые).
- Кремний откладывается в виде кремнезема в стенках растительных клеток, улучшая структурную жесткость и прочность клеточных стенок.
- Селен может повысить устойчивость растений к стрессу, вызванному ультрафиолетовым светом, замедлить биологическое старение и стимулировать рост.
Закон минимума Либиха
Хотя доступные количества каждого из основных питательных элементов важны, потребление питательных веществ растениями не является независимым по отдельности. Дефицит одного компонента может повлиять на общее усвоение питательных веществ растениями. В публикациях, выпущенных в 1840 и 1855 годах, немецкий химик-биолог Юстус фон Либих популяризировал открытие, получившее название «Закон минимума», который часто сокращается до закона Либиха. Этот Закон гласит, что рост растений контролируется не общим количеством доступных питательных веществ или ресурсов, а наличием самого дефицитного ресурса.Развитие растения ограничивается одним важным минералом, запасы которого относительно кратчайшие. Этот принцип используется для определения применения удобрений в современном сельском хозяйстве. Ключевым моментом является определение и внесение правильного питательного вещества, поскольку внесение неправильного удобрения не принесет почти никакой отдачи от урожая.
Поглощение растениями питательных веществ
Существует несколько способов передачи питательных веществ из почвы и других источников в растения.Некоторые растения, например те, которые питаются другими растениями или те, которые ловят насекомых, имеют специальные механизмы доступа к питательным веществам. Однако у большинства растений питательные вещества поглощаются растением через корни, в частности, через корневые волоски. Растение принимает растворенные питательные элементы через клеточную стенку.
Помимо прямого поглощения питательных веществ корнями, многие растения вступают во взаимовыгодные отношения с микроорганизмами. Некоторые растения, например бобовые, используют бактерии, такие как ризобии, для осуществления биологической фиксации азота, в результате чего атмосферный азот, который напрямую не доступен для растений, превращается в нитрат аммония, форму азота, которая доступна.Второй пример — взаимосвязь, которую образуют многие растения с грибами микоризы. Эта ассоциация обеспечивает гриб относительно постоянный и прямой доступ к источникам энергии, таким как сахар и крахмал, от растений. В свою очередь, длинные нитевидные структуры грибов действуют как продолжение корневой системы растения и увеличивают доступ растения к некоторым важным питательным веществам. Эта взаимосвязь дает растению гораздо больший доступ к питательным веществам, чем было бы в противном случае. В некоторых ситуациях основные макроэлементы, такие как фосфор, доступны растению только в количествах, необходимых для поддержания здорового роста благодаря этой связи.
Повышение плодородия почвы
До сих пор наше обсуждение было сосредоточено на питательных веществах почвы или химическом плодородии почвы, но это только один компонент плодородия. Под физическим плодородием понимаются физические свойства почвы, ее структура, текстура и водоудерживающие свойства, то, как вода течет к корням растений и как эти корни проникают в почву. Биологическое плодородие относится к организмам, живущим в почве, и их способности выполнять важные функции.Эти три компонента тесно взаимосвязаны, и даже оптимальные уровни питательных веществ могут иметь мало значения, если структура почвы и биология не способствуют здоровью растений.
Состав почвы, ее кислотность или щелочность, а также температура окружающей среды — это лишь некоторые из множества факторов, которые будут определять степень доступности питательных веществ для растений. Относительная важность этих факторов зависит от питательных веществ, растений и почвы. Наиболее важно то, что состав почвы влияет на то, насколько хорошо в ней удерживаются питательные вещества и вода.Глины и почвы, содержащие органические вещества, гораздо лучше удерживают питательные вещества и воду, чем песчаные почвы. Поскольку вода стекает с песчаных почв, она часто несет с собой питательные вещества. Это состояние называется выщелачиванием. Когда питательные вещества вымываются из верхнего слоя почвы, они становятся недоступными для использования растениями. Структура почвы также сильно влияет на микробиологию почвы. Растения, которые полагаются на популяции бактерий или грибов для поглощения питательных веществ, не будут расти, если почва не позволяет этим популяциям процветать.
Некоторые австралийские почвы, особенно в Западной Австралии, старые и выветрившиеся, а некоторые считаются старейшими почвами в мире. В течение долгого времени питательные вещества почвы вымывались, что приводило к низкому химическому плодородию почв. Например, в этих старых австралийских почвах уровень фосфора на 40% ниже, чем в почвах Соединенного Королевства, и на 50% ниже, чем в некоторых почвах Северной Америки. Однако важно отметить, что это сравнение неприменимо к более богатым сельскохозяйственным почвам восточной Австралии.До недавнего времени самым распространенным средством от дефицита питательных веществ было внесение удобрений. Однако сложность применения удобрений состоит в том, чтобы определить, какие питательные вещества действительно нуждаются в добавках и в каком количестве. Выбор времени также важен, поскольку потребности растений в питательных веществах меняются в зависимости от жизненного цикла растений. Влияние закона Либиха может замаскировать идентичность действительно дефицитных питательных веществ, поскольку симптомы дефицита не могут напрямую идентифицировать правильное дефицитное питательное вещество. Механизмы поглощения растениями, хотя они просто описаны выше, также могут быть сложными.Когда отсутствуют необходимые структурные и биологические условия, дополнительные удобрения не помогут или не помогут. Повышение жизнеспособности растений будет зависеть от улучшения структурного и биологического плодородия почвы. Неправильное внесение удобрений в лучшем случае является пустой тратой времени и денег, но также может иметь тяжелые экологические последствия.
Объем научных знаний, доступных австралийскому сельскохозяйственному производителю для точного анализа и интерпретации плодородия почвы, очень разнообразен.Возможность проверять и анализировать уровни питательных веществ в почве хорошо известна, и ее использование является финансово устойчивым и широко приемлемым для производителей. Много усилий было потрачено на картографирование, описание и классификацию австралийских почв на протяжении многих лет, особенно в сельскохозяйственных районах. Отдельным фермерам доступно тестирование почвы для анализа почвы с целью определения дефицита питательных веществ и токсичности. Исследования почвы могут быть дополнены анализом растений, который намного надежнее, чем диагностика визуальных симптомов, описанная выше.Однако возможность проводить такой же глубокий анализ структурного и биологического плодородия почвы появилась недавно и пока не имеет такого же уровня полезности во многих регионах. Анализ может быть дорогостоящим и в настоящее время менее широко используется сельскохозяйственным сообществом. Конечная цель всестороннего понимания всех трех компонентов плодородия почвы в сочетании с одинаково всесторонним пониманием того, как эти компоненты взаимосвязаны друг с другом, а также с водой и углеродом в сельскохозяйственных экосистемах, вероятно, останется достойной целью в обозримом будущем.
Заключение
Обманчивая простота концепции, согласно которой растениям нужны питательные вещества для роста и процветания, возможно, привела к недооценке глубинной сложности метаболического процесса, вовлеченного в перенос основных химических элементов из почвы в ткань растений. Мы все в какой-то степени осознаем необходимость того, чтобы формы жизни, включая нас самих, получали питание из окружающей среды, чтобы жить, расти и воспроизводиться.Несмотря на эту простоту, важные минеральные вещества, которые растения получают из почвы, являются лишь одним взаимосвязанным компонентом в устрашающе сложной системе взаимосвязей и циклов. Задача улучшить наше понимание этих отношений и извлечь из них практические и устойчивые методы ведения сельского хозяйства вполне может укрепить нашу способность поддерживать сельскохозяйственное производство в будущем.
*****
Любые мнения или взгляды, выраженные в этом документе, принадлежат отдельному автору, если не указано, что они принадлежат Future Directions International.
Опубликовано Future Directions International Pty Ltd.
80 Birdwood Parade, Далкейт, Вашингтон, 6009, Австралия.
Тел .: +61 8 9389 9831 Факс: +61 8 9389 8803
Электронная почта: [адрес электронной почты защищен] Веб-сайт: www.futuredirections.org.au
Питательные вещества для растений в почве
Почва — основной источник питательных веществ, необходимых растениям для роста. Три основных питательных вещества — это азот (N), фосфор (P) и калий (K).Вместе они составляют трио, известное как NPK. Другими важными питательными веществами являются кальций, магний и сера. Растения также нуждаются в небольшом количестве железа, марганца, цинка, меди, бора и молибдена, известных как микроэлементы, потому что растениям нужны только следы. Эти питательные вещества играют сложную роль в росте растений, и в этом документе приводится лишь краткое описание.
Основные элементы
Азот (N)
Азот — ключевой элемент в росте растений. Он содержится во всех растительных клетках, в растительных белках и гормонах, а также в хлорофилле.
Атмосферный азот является источником азота почвы. Некоторые растения, такие как бобовые, фиксируют атмосферный азот в своих корнях; в противном случае предприятия по производству удобрений используют азот из воздуха для производства сульфата аммония, нитрата аммония и мочевины. При внесении в почву азот превращается в минеральную форму, нитрат, так что растения могут усваивать его.
Почвы с высоким содержанием органических веществ, такие как шоколадные почвы, обычно содержат больше азота, чем подзолистые почвы. Нитраты легко вымываются из почвы сильными дождями, что приводит к ее подкислению.Вам нужно часто вносить азот в небольших количествах, чтобы растения использовали его весь, или в органической форме, такой как компостированный навоз, чтобы уменьшить вымывание.
Фосфор (P)
Фосфор помогает передавать энергию солнечного света растениям, стимулирует ранний рост корней и растений и ускоряет созревание.
Очень немногие австралийские почвы имеют достаточно фосфора для устойчивого выращивания сельскохозяйственных культур и пастбищ, и Северное побережье не является исключением. Самый распространенный источник фосфора на Северном побережье — суперфосфат, сделанный из каменного фосфата и серной кислоты.Все удобрения содержат фосфор; навоз животных, выращиваемых на зерно, является особенно богатым источником.
Калий (K)
Калий повышает жизнеспособность и устойчивость растений к болезням, помогает формировать и перемещать крахмал, сахар и масла в растениях и может улучшить качество плодов.
На многих песчаных почвах Северного побережья калия мало или его не хватает. Кроме того, сильное удаление калия может происходить на почвах, используемых для интенсивного выпаса скота и интенсивных садовых культур (таких как бананы и заварные яблоки).
Муриат калия и сульфат калия являются наиболее распространенными источниками калия.
Кальций (Ca)
Кальций необходим для здоровья корней, роста новых корней и корневых волосков, а также для развития листьев. Обычно его не хватает на кислых почвах Северного побережья. Известь, гипс, доломит и суперфосфат (смесь фосфата кальция и сульфата кальция) поставляют кальций. Лайм — самый дешевый и подходящий вариант для Северного побережья; доломит полезен при дефиците магния и кальция, но если его использовать в течение длительного периода, соотношение кальция / магния нарушится.Суперфосфат полезен там, где необходимы кальций и фосфор.
Магний (Mg)
Магний является ключевым компонентом хлорофилла, зеленого красящего вещества растений, и жизненно важен для фотосинтеза (преобразования солнечной энергии в пищу для растений). Дефицит встречается в основном на песчаных кислых почвах в районах с большим количеством осадков, особенно если они используются для интенсивного садоводства или молочного животноводства. Интенсивное внесение калия в удобрения также может вызвать дефицит магния, поэтому производители бананов должны следить за уровнем магния, потому что бананы являются большими потребителями калия.
Дефицит магния можно преодолеть с помощью доломита (смесь карбоната магния и кальция), магнезита (оксида магния) или солей эпсома (сульфата магния).
Сера (S)
Сера входит в состав аминокислот в растительных белках и участвует в процессах производства энергии в растениях. Он отвечает за многие компоненты вкуса и запаха растений, такие как аромат лука и капусты.
Дефицит серы не является проблемой для почв с высоким содержанием органических веществ, но она легко выщелачивается.На Северном побережье морские брызги являются основным источником атмосферной серы. Суперфосфат, гипс, элементарная сера и сульфат аммиака являются основными источниками удобрений.
Микроэлементы
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.
Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie. - Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г.,
браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере. - Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.
Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.