РОЛЬ ЛИПИДОВ В ПИТАНИИ. «БИОЛОГИЧЕСКАЯ ХИМИЯ», Березов Т.Т., Коровкин Б.Ф.
РОЛЬ ЛИПИДОВ В ПИТАНИИ
Липиды
являются обязательной составной частью сбалансированного пищевого рациона
человека. В среднем в организм взрослого человека с пищей ежесуточно поступает
60–80 г жиров животного и растительного происхождения. В пожилом возрасте, а
также при малой физической нагрузке потребность в жирах снижается, в условиях
холодного климата и при тяжелой физической работе – увеличивается.
Значение
жиров как пищевого продукта весьма многообразно. Жиры в питании человека прежде
всего имеют важное энергетическое значение. Энергетическая ценность жиров выше,
чем белков и углеводов. Известно, что при окислении 1 г жиров организм получает
38,9 кДж (9,3 ккал), тогда к а к при окислении 1 г белков или углеводов – 1 7 ,
2 кДж ( 4 , 1 ккал). Кроме того, жиры являются растворителями витаминов A, D, Е
и К, в связи с чем обеспеченность организма этими витаминами в значительной
степени зависит от поступления жиров в составе пищи. С жирами в организм
вводятся и некоторые полиненасыщенные жирные кислоты (линолевая, линоленовая,
арахидоновая), которые относят к категории незаменимых (эссенциальных) жирных
кислот, так как ткани человека и ряда животных потеряли способность
синтезировать их. Эти кислоты условно объединены в группу под названием
«витамин F».
Известно также,
что жир обеспечивает вкусовые качества пищи; кроме того, он необходим для ее
приготовления и хранения. Все это привело к тому, что потребление жира в
высокоразвитых странах столь велико, что за его счет покрывается более 35%, а
во многих странах более 40% энерготрат организма. Это в свою очередь очень
часто ведет к тому, что прием обогащенной жирами пищи перекрывает
физиологические потребности организма в энергии. Отсюда такие неблагоприятные
явления, как ожирение значительной части населения. Поэтому знание метаболизма
липидов нормального организма необходимо и для понимания причин многих
болезней. Известно, что нарушения метаболизма липидов возникают, например, как
при избыточном, так и при недостаточном приеме жиров, дефиците тех или иных
ферментов, при дисбалансе гормонов и т.д.
Предыдущая страница |
Следующая страница
СОДЕРЖАНИЕ
Липиды. Виды, значение, пищевая ценность липидов
Содержание страницы
Липиды – необходимые компоненты пищи, которые в сочетании с другими полимерами биопищевого сырья и продуктов обеспечивают нормальную жизнедеятельность организма и здоровье человека. Вместе с тем роль пищевых липидов в технологии продуктов питания и пищи трудно переоценить. Они достаточно распространены в природе, в том числе источниках пищи животного, растительного и микробного происхождения.
В пищевых источниках липиды присутствуют в протоплазме клеток и мембранах, а также образуют специализированные ткани – резервный жир. Протоплазматический и резервный жиры составляют основную часть липидов (примерно 10–20 % массы тела). Протоплазматический (конститутивный) жир входит в состав всех структур клеток органов и тканей и в течение всей жизни остается практически на одном уровне – около 25 % общего жира организма.
Резервный жир запасается в организме, и его количество меняется в зависимости от возраста, пола, условий питания, характера деятельности. Это обстоятельство широко используется при составлении рационов и диет на основе глубоких исследований структуры, функций и метаболизма липидов.
В настоящее время установлено, что уровень потребления липидов, как и белков, должен в среднем составлять 80–100 г/сут. Современный уровень исследований позволил выработать определенные требования и к качественному составу липидов, поскольку многие из них могут способствовать развитию заболеваний или, наоборот, служить профилактическим и даже лечебным средством. Липиды обладают и функциональнотехнологическими свойствами, в частности способностью образовывать эмульсии – основу большого числа пищевых продуктов. Достижения химии липидов позволяют создавать пищевые добавки на основе природных липидов, улучшающих консистенцию и повышающих биологическую ценность продуктов.
Наличие липидов в каждом виде тканей различных организмов говорит о важной роли этих веществ в жизнедеятельности организма. Наиболее изучены пластические и энергетические функции липидов. Первое связано с тем, что они являются частью структуры многих важных соединений – гормонов, сложных белков, витаминов, составляя главные компоненты клеточных мембран – «контейнеров» для протекания основных метаболических процессов.
Мембраны образуют клеточный покров, отделяюший содержимое клетки от окружающей среды, обеспечивают пространственное разделение метаболических процессов внутри клетки. С наличием липидов в мембранах связывают работу многочисленных ферментных и транспортных систем, распознавание участков, обеспечивших связывание определенных гормонов и восприятие иных сигналов из внешнего окружения.
Энергетическая функция липидов подтверждается тем, что они обеспечивают примерно 25–30 % всей энергии организма. При полном сгорании 1 г липидов выделяется более чем в два раза больше энергии по сравнению с белками и углеводами. Запасные (резервные) липиды играют роль энергетического депо и локализуются в виде специализированных тканей (подкожная клетчатка, сальник, околопочечная капсула и т. п.). Они принимают участие в процессах терморегуляции, предохраняют кожу от высыхания, защищают органы от сотрясения, обеспечивают всасывание из кишечника жирорастворимых витаминов, являются потенциальным резервом эндогенной воды в организме (при окислении 100 г жира образуется 100 см3 воды) и др.
В настоящее время установлено, что некоторые липиды обладают биологической активностью. Многие из них связаны со структурной и функциональной организацией нервной системы, органов кроветворения и коммуникационных систем высших организмов.
1. Классификация липидов
Современная классификация липидов основывается на комплексе их свойств: химической природе, биологических функциях, способности к растворению при взаимодействии с некоторыми реагентами, например щелочами. В целом к липидам относят природные органические соединения, нерастворимые в воде, но растворимые в органических растворителях (бензол, бензин, петролейный эфир, этиловый эфир, ацетон, хлороформ, сероуглерод, метиловый и этиловый эфиры и т. п.). Пищевые липиды легко утилизируются животными организмами.
Класс липидов подразделяют на жиры и жироподобные вещества. Одна из главных групп липидов обозначается общим термином – жиры, а остальные сходные с жирами по растворимости соединения, входящие в класс липидов, составляют группу липоидов (жироподобных веществ).
По химической структуре липиды делят на простые и сложные.
Некоторые авторы выделяют еще и производные липидов.
Простые липиды не содержат азота, фосфора и серы. К ним относят нейтральные липиды, являющиеся производными высших жирных кислот, одно-, двух- и многоатомных спиртов, альдегидов, а также их структурные компоненты (спирты, карбоновые кислоты). Таким образом, простые липиды состоят из эфиров жирных кислот и глицерина, а также высших или полициклических спиртов.
Сложные липиды имеют многокомпонентные молекулы, составные части которых соединены химическими связями различного типа и имеют разнообразное химическое строение. К ним относят фосфолипиды и гликолипиды.
Фосфолипиды – разнообразная и важная группа сложных липидов. Их структура образована фосфорной кислотой, насыщенными и ненасыщенными жирными кислотами, альдегидами, спиртами, азотистыми основаниями (холин, этанололамин), аминокислотами, связанными между собой сложной или простой эфирной или амидной связью.
Гликолипиды наряду с многокомпонентным спиртом и высшей жирной кислотой имеют в своем составе углеводы. Необходимо отметить, что некоторые гликолипиды могут быть отнесены также к простым липидам. В последнее время установлено, что гликолипиды присутствуют не только в нервных тканях, но и обнаружены в хлоропластах растений.
По функциям в организме липиды разделяют на структурные, запасные и защитные.
К структурным липидам относят фосфолипиды, которые, образуя сложные комплексы с белками (липопротеины) и углеводами, участвуют в разнообразных процессах живой клетки. К ним также относят глико-, сульфолипиды и некоторые другие.
Запасные липиды образуются в основном ацилглицеринами, которые в растениях накапливаются большей частью в плодах и семенах, а у животных и рыб – в подкожных тканях и тканях, окружающих внутренние органы, а также печени, мозговой и нервной тканях.
Содержание их зависит от многих факторов (вида, возраста, питания и т. п.) и в отдельных случаях составляет 95–97 % всех выделяемых липидов. Защитные липиды – воски и их производные – у растений покрывают поверхность листьев, семян и плодов, предохраняя их от воздействия внешних факторов. Значительное количество разнообразных липидов содержится в молоке животных (в %): дельфин – 46,0, кит – 45,0, слон – 20,0, коза – 4,8, корова – 3,6.
Известна классификация липидов по их способности реагировать со щелочами. По отношению к щелочам их делят на две большие группы: омыляемые и неомыляемые. К группе омыляемых липидов относят простые и сложные липиды, которые при взаимодействии со щелочами гидролизуются с образованием солей высокомолекулярных кислот, получивших название «мыла».
В суммарной фракции природных липидов идентифицированы вещества, которые обладают такой же растворимостью, как и липиды, но не способны омылиться. Состав фракции представлен свободными высшими жирными кислотами, высшими и полициклическими спиртами (стеролами), их производными (стероидами), жирорастворимыми витаминами, высшими гомологами предельных и непредельных углеводородов и другими соединениями.
Липиды являются важными компонентами пищи, во многом определяют ее пищевую ценность и вкусовые качества. Исключительно велика роль липидов в разнообразных процессах пищевой технологии. Порча зерна и продуктов его переработки при хранении связана с изменением липидного состава входящего комплекса.
Липиды, выделенные из растений и животных, – важнейшее сырье для получения пищевых и технических продуктов: растительное масло, животные жиры, в том числе сливочное масло, маргарин, глицерин, жирные кислоты.
2. Физиологическое значение, пищевая и биологическая ценность липидов
Важное структурное и функциональное значение липидов в организме делает их необходимыми компонентами пищи, которые должны присутствовать в рационах в определенных количествах и соотношениях с другими пищевыми веществами. Учитывая важное физиологическое значение и эссенциальность отдельных представителей липидов в обеспечении здорового питания, весьма важны их качественный состав и химическая природа.
Необходимым свойством пищевых липидов является их переваримость и усвояемость.
Основными факторами переваривания жиров считаются: специфические ферменты – липазы; наличие оптимальных условий проявления их активности; наличие эмульгаторов – веществ, понижающих поверхностное натяжение и предупреждающих склеивание частиц жира. Частично такие веществa содержатся в природных маслах и жирах.
Липазы способны гидролизовать жиры только в эмульгированном состоянии. В организме основными эмульгаторами жира являются соли желчных кислот и сывороточный альбумин. Пищеварительный канал – начальный путь превращения липидов в организме.
В ротовой полости отсутствуют условия переваривания жира, в связи с чем заметных изменений в жировых фракциях не наблюдается. В желудке присутствует липаза, но из-за сильной кислой внутренней среды желудка активность липазы невелика, т. к. рНоптимум ее действия находится в слабощелочной среде. Однако природные тонкодиспергированные (эмульгированные) жиры (молока, яичного желтка) в желудке расщепляются.
Вместе с тем желудок в переваривании жиров имеет большое значение: он регулирует поступление жиров в кишечник. Кроме того, протеолитические ферменты желудка (пепсин, ренин) переваривают (гидролизуют) липопротеиновые комплексы пищи, обеспечивая доступность жировых фрагментов для переваривания в кишечнике.
В кишечнике имеются необходимые условия для эффективного расщепления жиров под действием ферментов. Именно поэтому тонкий кишечник является основным местом переваривания жиров. Соки поджелудочной железы и кишечника богаты липолитическими ферментами, к тому же слабощелочная реакция среды (рН 7,8–8,0) идеально обеспечивает эффективность их действия на субстраты. Эмульгирование жира происходит под действием двух основных факторов.
Во-первых, в просвет кишечника поступают с желчью желчные кислоты, обладающие известным эмульгирующим действием и активирующие пищеварительную липазу; они синтезируются и секретизируются поджелудочной железой и в кишечнике.
Во-вторых, важную роль выполняет белковый кофактор – колипаза, которая связывает липазу в присутствии желчных кислот, в результате чего рН-оптимум действия липазы смещается с 9 до 6.
Липазы поджелудочной железы и кишечника обладают позиционной специфичностью, т. е. разрывают сложноэфирные связи не все сразу, а по одной в соответствии со специфичностью действия. Дополнительно диспергирование жира обеспечивается перистальтикой кишечника и постоянно выделяющимся углекислым газом, который образуется при нейтрализации кислого содержимого желудка под действием бикарбонатов кишечника. Газ обеспечивает высокую дисперсность пищевых масс, в том числе жиров.
Итак, в тонком кишечнике в основном нейтральные жиры расцепляются на составные фрагменты – глицерин и высшие жирные кислоты. Часть жиров гидролизуется до ди- и моноглицеринов. Непереваренный жир поступает в толстый кишечник и выделяется с калом.
Процесс всасывания продуктов гидролизa жиров довольно сложен в случае жирных кислот, т. к. глицерин водорастворим и свободно всасывается и поступает в кровь. Жирные кислоты переходят в растворимое состояние путем предварительного образования холиновых комплексов, представляющих собой соединения 2–4 парных желчных кислот и одной жирной кислоты. Все желчные кислоты (стероидные монокарбоновые оксикислоты) образуются в печени из холестерина. Они (главным образом холевая и дезоксихолевая кислоты) способны взаимодействовать с гликоколом (глицином) или таурином с образованием пептидоподобной связи. Такие соединения называют парными желчными кислотами.
Холиновые комплексы растворимы в воде и свободно всасываются в стенку кишечника, где вновь распадаются до составных частей. При этом желчные кислоты поступают в воротную вену и попадают в печень, а затем в желчный пузырь и в составе желчи вновь поступают в кишечник. Таким образом, за счет кругооборота небольшое количество желчных кислот обеспечивает переваривание и всасывание значительных количеств жирных кислот.
Неполностью расщепленные или совсем нерасщепленные жиры также могут всасываться в кишечнике, если они тонко эмульгированы и размер их частиц не превышает 0,5 нм.
В итоге в стенке кишечника накапливаются глицерин и жирные кислоты, что обеспечивает возможность первичного синтеза жиров, но уже специфичных для данного организма.
Холестерин пищи содержится как в свободной форме, так и в виде стероидов, которые подвергаются гидролизу до составных частей. Холестерин, подобно жирным кислотам, всасывается с помощью парных желчных кислот.
Накопившиеся в процессе пищеварения и всасывания глицерин, жирные кислоты, холестерин, фосфорная кислота, азотистые основания, синтезированные моно-, ди- и триглицериды, фосфолипиды, эфиры холестерина в стенке кишечника образуют хиломикроны – транспортные формы жира, структурная схема которых показана на рис. 1.
Рис. 1. Структурные компоненты хиломикрона
В составе хиломикронов основная часть жирорастворимых соединений поступает в лимфатическую систему, а затем в кровь.
После приема пищи содержание жира в крови повышается, в результате чего примерно через 3 ч она мутнеет, а через 4–6 ч постепенно осветляется за счет действия фермента липопротеинлипазы, которая вызывает распад хиломикронов на более мелкие части, а триглицеридов – до жирных кислот и глицерина. Они переносятся к различным органам и тканям, где подвергаются специфическим превращениям.
Значительная часть жира поступает в печень, а меньшая часть резервируется в жировых депо. В мышцы, сердце поступают в основном комплексы жирных кислот, глицерина и белков, которые там распадаются, освобождая важные для этих органов жирные кислоты.
Из печени жиры в виде гликопротеинов распространяются по всему организму, где и используются по мере необходимости.
Поступивший в клетки нейтральный жир под действием тканевых липаз гидролизуется с распадом на глицерин и жирные кислоты. При этом глицерин активируется при участии специфического фермента глицерокиназы и АТФ, а затем превращается в фосфоглицериновый альдегид и включается в углеводный обмен, распадаясь до молочной кислоты или СО2 и Н2О, а также участвует в синтезе гликогена, образовании глицерина путем, обратным его распаду. Жирные кислоты подвергаются β-окислению, которое начинается с их активации при участии АТФ, кофермента А (КоА), витаминов С и В2 и фермента тиокиназы.
Наиболее активно окисление жирных кислот протекает в печени, мышцах, жировых депо, сердце и в меньшей степени – в других органах.
Ненасыщенные жирные кислоты имеют несколько иной путь окисления с участием специального набора ферментов.
Наряду с распадом жиров в клетках постоянно происходит липогенез – синтез специфического для каждого органа нейтрального жира. Процесс включает образование глицерина, жирных кислот с последующим синтезом триацилглицеринов. При этом синтез глицерина возможен из продуктов распада углеводов и жиров.
Именно в этом усматривается тесная связь успешного углеводного и жирового обменов.
Ненасыщенные жирные кислоты не синтезируются в организме или синтезируются ограниченно, что требует их постоянного поступления с продуктами питания.
В тканях постоянно происходит также обмен фосфолипидов. Под действием тканевых фосфолипаз они гидролизуются на составные части. Глицерин и жирные кислоты превращаются вышеописанными способами.
Фосфорная кислота используется в неизменном виде, в то время как азотистые основания превращаются специфически. При этом серин, например, участвует в биосинтезе белков и обмене некоторых других веществ. Холин (как витаминоподобное вещество) незаменим для обмена веществ, служит для синтеза ацетилхолина – одного из медиаторов нервной системы, а также участвует в синтезе фосфолипидов, препятствуя повышенному накоплению жира в печени.
В организме также происходит постоянный синтез липоидов, в котором участвуют глицерофосфат, активированные жирные кислоты, фосфорная кислота, активированные азотистые основания.
Холестерин в организме подвергается различным превращениям и служит источником важнейших соединений (гормонов, витамина D3, желчных кислот). В клетках печени, мозга, других органов холестерин постоянно синтезируется в количестве 0,8–1,5 г/сут. Установлено, что синтез холестерина начинается с конденсации трех молекул ацетил-КоА и образования промежуточного продукта – мевалоновой кислоты, а затем образуется циклическая форма – сквален, который затем превращается в холестерин.
Процессы обмена жира в организме регулируются нейрогуморальным путем, т. е. при участии нервной системы всех уровней и гормонально. Большое значение имеют гормоны инсулин, адреналин, тироксин и гормон роста. Важными регуляторными механизмами обмена жиров являются физиологические и внешние факторы: пол, возраст, характер профессиональной деятельности, режим дня.
Чрезвычайное значение имеет питание. Патологии при нарушениях обмена жиров могут быть наследственной природы или приобретаются в результате неправильного питания и нездорового образа жизни. Так, нерегулярное и несбалансированное питание, избыточность углеводной пищи, малоподвижный образ жизни и труда способствуют отложению повышенного количества жира, развитию ожирения и ряда сопутствующих заболеваний.
Очевидно, что за счет питания можно достичь хорошего эффекта профилактики и лечения заболеваний. Давая общую биологическую оценку жировым пищевым продуктам, следует подчеркнуть, что она определяется тремя основными факторами – количеством и качеством жирорастворимых витаминов, полиненасыщенных жирных кислот и фосфатидов.
Однако ни один природный жир, используемый в питании, не является полноценным во всех отношениях. Биологическая полноценность пищевых жиров может быть достигнута сочетанием продуктов в рационах за счет использования растительных и животных источников. Оптимальное сочетание достигается при использовании 70–80 % (примерно 2/3) растительных и 20–30 % (около 1/3) животных жировых продуктов. Кроме этого в нормально сбалансированном рационе питания взрослого человека должно выдерживаться соотношение белков, жиров и углеводов 1:1:4. В пожилом возрасте, а также при малой физической нагрузке и занятости умственным трудом потребность в жире снижается.
Недостаточное поступление жира с пищей вызывает разнообразные нарушения обменных процессов, а также становится причиной развития авитаминозов жирорастворимых витаминов (поскольку для их всасывания необходим жир) и снижения количества эссенциальных жирных кислот.
Дисбаланс может возникать при нарушении переваривания и всасывания липидов по причине недостаточного синтеза липолитических ферментов в пищеварительном тракте и секреции желчи. Нарушения жирового обмена отмечаются и при недостатке в организме липотропных веществ типа холина, метионина, витамина F (фактора F), которые предохраняют печень от повышенного отложения жира путем участия в синтезе фосфолипидов.
Следствием нарушения жирового обмена является развитие ожирения – болезни современной цивилизации. Сегодня в большинстве развитых стран более 50 % населения имеет избыточную массу тела. Ожирение – патология, характеризующаяся повышенным отложением жира во всем организме.
Значительный интерес представляют нарушения обмена холестерина. Причиной гипохолестеринемии является нарушение равновесия между количеством распавшегося и синтезированного холестерина в организме. С пищей при нормальной диете его поступает около 0,2–0,5 г/сут, и это количество практически не влияет на уровень холестерина в организме. Считают, что основная роль принадлежит эндогенному холестерину, количество которого в организме достигает 0,8–1,5 г/сут. Отмечено, что увеличение его содержания наблюдается при избыточном потреблении жиров и углеводов и нарушении процессов использования ацетил-КоА. При избытке холестерина развиваются различные атеросклерозы.
Лечение направлено на нормализацию жирового обмена и торможение эндогенного синтеза холестерина в организме. Вместе с тем безусловным фактором профилактики атеросклерозов выступают правильное питание и дозированное употребление в пищу продуктов с высоким содержанием холестерина и жиров. По этой причине большинство современных продуктов снабжены информацией о массовой доле холестерина.
В настоящее время установлено, что общее содержание жиров в рационе рекомендуется поддерживать на уровне 30–35 % энергетической ценности рациона, в массовом выражении это в среднем 80–100 %. Высокое содержание животных жиров нежелательно, поскольку при избытке насыщенных жирных кислот нарушается обмен липидов. Повышается уровень холестерина в крови, увеличивается риск развития атеросклероза, ожирения, желчнокаменной болезни.
Важно иметь информацию о качественном составе ненасыщенных жирных кислот в липидах (например, о содержании моно- и полиненасыщенных жирных кислот). К тому же полиненасыщенные жирные кислоты (ПНЖК), не синтезируемые в организме человека, относят к незаменимым (линолевая, линоленовая, арахидоновая). Арахидоновая кислота образуется в организме из линолевой при участии витамина В6. Линолевая образует и другие ПНЖК. По расположению первой двойной связи относительно метильной группы в молекуле ПНЖК относят к группам омега-6 (линолевая, γ-линоленовая, арахидоновая) и омега-3 (α-линоленовая, эйкозапентаеновая, докозагексаеновая).
Установлена четкая обратная взаимосвязь между суточным потреблением омега-3 жирных кислот и степенью атеросклеротических поражений коронарных сосудов. При этом, чем больше омега-3 содержится в тканях организма, тем меньше проявлений атеросклероза. Жирные кислоты омега-3 снижают уровень триглицеридов в сыворотке крови, уменьшают риск образования тромбов в сосудах, способствуют синтезу простагландинов, поддерживающих иммунный статус организма и необходимый для нормальной функции надпочечников и щитовидной железы.
Кислоты омега-3 содержатся в жире рыбы, льняном и соевом маслах, в масле грецкого ореха; источником жиров омега-6 служат подсолнечное, кукурузное и сафлоровое масла. Очень важно поддерживать оптимальный уровень соотношения между омега-3 и омега-6 жирными кислотами. Рекомендуемое соотношение в рационе омега-6:омега-3 составляет для здорового человека 4:1, для лечебного питания – от 3:1 до 2:1.
Среднее содержание ПНЖК в рационе в пересчете на линолевую кислоту должно составлять 4–6 % общей калорийности пищи. Избыток ПНЖК, как и недостаток, отрицательно сказывается на здоровье человека.
Большое значение в питании имеют и фосфолипиды, потому что они также входят в состав мембран клеток человеческого организма и участвуют в липидном обмене, способствуя перевариванию и правильному обмену жиров, в частности их транспорту из печени в другие участки организма. Недостаток фосфатидов в рационе приводит к накоплению жира в печени, к ее ожирению, а затем и циррозу. Фосфатиды участвуют в регуляции обмена веществ, построении структуры нервной ткани, клеток печени, мозга; снижают уровень холестерина в крови и препятствуют его отложению в стенках кровеносных сосудов, что является важнейшей профилактической мерой в отношении развития атеросклероза и сопутствующих ему сердечнососудистых заболеваниях. Суточная потребность здорового взрослого человека в фосфатидах составляет 5–10 г.
Наиболее распространенный в пищевых продуктах фосфолипид – лецитин, содержание которого составляет (в %): в яйцах – 3,4; нерафинированном растительном масле 1,0–2,0; сырах – 0,5–1,1; мясе – 0,8; птице – 0,5–2,5; в зерне и бобовых 0,3–0,9. Он присутствует во всех клетках человеческого организма. За счет содержания метильных групп активно проявляет себя в липидном обмене, в частности предотвращает жировое перерождение печени, обеспечивает нормальное функционирование нервной ткани, повышает умственную работособность, положительно влияет на сексуальную активность, способствует снижению уровня холестерина в крови.
Следует придерживаться рекомендаций по ограничению содержания холестерина в рационах – в среднем не более 500 мг в сутки. Больше всего холестерина содержится (в %): в яйцах – 0,57; сливочном масле – 0,17–0,27; печени – 0,13–0,27; мясе – 0,06–1,00; рыбе – до 0,30. Вместе с тем характеристика основных пищевых липидов требует присутствия растительных и животных жиров в рационах, т. к. они в равной степени необходимы человеку. Животные жиры – единственный источник витаминов А и D, растительные – витаминов Е и β-каротина.
К жировым продуктам растительного происхождения относят масла, на 95–97 % состоящие из триацилглицеринов и небольшого количества сопутствующих веществ (фосфатидов, токоферолов, стероидов, пигментов). Масла получают из семян и плодов высокомасличных культурных растений (подсолнечное, хлопковое, льняное, конопляное, соевое, оливковое, кукурузное и другие масла). Сопутствующие вещества масла повышают его биологическую ценность, стойкость при хранении, обусловливают ароматические и вкусовые особенности, окраску. Их содержание колеблется в зависимости от природных факторов, условий выращивания, уборки, хранения, способов тепловой обработки и принятой технологии в целом.
Масличные семена и продукты их переработки (кроме масла и белка, являющегося побочным продуктом переработки) содержат богатейший комплекс биологически активных соединений, в том числе витаминной и провитаминной природы. Они исключительно богаты жирорастворимыми витаминами и провитаминами – токоферолами, стероидами и каротиноидами, в их составе много водорастворимых витаминов – тиамина, рибофлавина, пиридоксина, биотина, фолиевой кислоты, пантотеновой кислоты, инозита, аскорбиновой кислоты.
Разнообразен фосфолипидный комплекс масличных семян – в его составе фосфатидил-холины, фосфатидилэтаноламины, фосфатидилсерины, инозитфосфатиды, фосфатидные кислоты и их соли. К тому же растительные источники жиров содержат уникальный набор макро-, микро- и ультрамикроэлементов, суммарное содержание которых почти в два раза превышает их количество в семенах других культур. Фосфолипиды – важный компонент комплекса липидов растений.
Они отличаются от триацилглицеринов тем, что состав жирных кислот характеризуется большей степенью ненасыщенности.
Гликолипиды в растениях достаточно разнообразны, однако детально не изучены.
Гликолипиды растений имеют выраженную полярность. Гидрофобная часть молекул образована жирными кислотами с большим числом атомов углерода.
К неомыляемым липидам растительного происхождения относят соединения двух типов: стероиды и терпены. Они имеют одинаковые структурные фрагменты – пятиуглеродные блоки, в них также обнаруживаются спирты витаминов A, D, Е.
Каротиноиды и хлорофиллы – наиболее изученные соединения, определяющие окраску природных масел; они являются производными изопрена или содержат в своем составе изопреновые группировки.
Группа каратиноидов представлена 65–75 природными пигментами, которые имеют цвет от светло-желтого до темно-красного. Они подразделяются на углеводороды и спирты. Углеводороды – каротины С40Н56 – находятся в растительных маслах в виде смеси α-, β- и γкаротинов.
Хлорофиллы высших растений представляют собой смесь двух близких по строению соединений: хлорофилла α (C55Н72О5N4Mg) – сине-черного вещества с температурой плавления 117–120 °С и хлорофилла β (C55H70O6N4Mg) – желто-зеленого вещества с температурой плавления 86–92 °С. Соотношение их в растительных тканях 3:1.
Хлорофиллы – воскообразные вещества, хорошо растворимые в ацетоне, спирте и растительных маслах. Растворимость хлорофиллов в растительных маслах и органических растворителях обусловлена тем, что хлорофилл имеет длинную гидрофобную терпеноидную боковую цепь, которая представляет собой остаток спирта фитола, связанный сложноэфирной связью с остатком пропионовой кислоты.
Недозревшие масличные семена содержат значительное количество хлорофиллов. Затем, по мере созревания семян, хлорофиллы исчезают и в зрелых семенах присутствуют в виде следов. Присутствие хлорофилла и его видоизменений в растительных маслах следует считать нежелательным. Установлено, что присутствие хлорофилла инициирует окислительные процессы, и масло оказывается нестойким при хранении.
На предприятиях общественного питания и пищевой промышленности широко применяется технологический прием тепловой обработки изделий из мяса, рыбы, овощей, теста – фритюрная жарка. С его помощью получают вкусную, калорийную, с привлекательным внешним видом продукцию, которая пользуется большим спросом у потребителя.
Однако при фритюрной жарке жиры подвергаются довольно жесткому воздействию: они нагреваются при высоких температурах (160–190 °С) в течение нескольких часов, иногда дней; к поверхности горячего жира, как правило, имеется свободный доступ воздуха; через слой жира постоянно проходит влага, выделяемая продуктом, при этом жир пенится, что увеличивает поверхность соприкосновения его с воздухом; в жир попадают частицы обжариваемого продукта и, если нет постоянной фильтрации жира или «холодной» зоны у аппарата, обугливаются и загрязняют жир продуктами пирогенетического распада содержащихся в них органических веществ. Такие условия нагрева вызывают глубокие изменения в жирах.
При термическом воздействии в жирах проходят четыре основных процесса: окисление, полимеризация, гидролиз и декструкция. В маслах, богатых полиненасыщенными жирными кислотами, процессам окисления и полимеризации предшествует (а затем и сопутствует) процесс конъюгирования двойных связей (рис. 2).
Рис. 2. Схема процессов, происходящих при гидротермическом воздействии на жиры
На схеме (рис. 4) видно, что это не параллельно протекающие, а взаимосвязанные, переплетающиеся процессы.
Просмотров:
153
Пищевая ценность липидов
Пищевые
жиры и масла являются обязательным
компонентом пищи, источником энергетического
и пластического материала для человека,
поставщиком необходимых веществ, таких
как: ненасыщенные жирные кислоты,
фосфолипиды, жирорастворимые витамины,
стерины. Рекомендуемое содержание жиров
в рационе человека по калорийности
составляет 30 – 33 % или 90 – 107 г в сутки.
Среднем считается норма в 102 г в сутки.
В питании имеет значение не только
количество, но и химический состав
жиров. Линолевая и линоленовые кислоты
не синтезируются в организме человека,
арахидоновая кислота синтезируется из
линолевой кислоты при участии витамина
В6.
Поэтому они получили название незаменимых
или эсенциальных
жирных кислот.
В последние годы часто употребляется
термин «полиненасыщенные жирные кислоты
семейства омега – 3», в эту группу входят
ά – линоленовая, эйкозапентаеновая,
докозагексаеновая кислоты, содержащие
несколько кратных связей и «полиненасыщенные
жирные кислоты семейства омега – 6», в
эту группу вхадит арахидоновая кислота.
Ненасыщенные
жирные кислоты участвуют в расщеплении
липопротеидов, холестерина, предотвращают
образование тромбов, снижают воспалительные
процессы.
Липиды оказывают
влияние на обмен веществ в клетках,
входят в состав клеточных мембран,
влияют на кровяное давление, выводят
из организма холестерин, при этом
повышается эластичность стенок
кровеносных сосудов. Повышенной
биологической активностью обладают
арахидоновая и линолевые кислоты. Среди
продуктов питания наиболее богаты
полиненасыщенными жирными кислотами
растительные масла. Арахидоновая кислота
содержится в яйцах, субпродуктах.
Сбалансированный состав ежедневного
рациона человека должен содержать 10 –
20 % полиненасыщенных жирных кислот, 50 –
60 % мононенасыщенных жирных кислот, 30 %
насыщенных жирных кислот. \это
обеспечивается при использовании в
рационе одной трети растительных и двух
третей животных жиров.
Фосфолипиды
участвуют в построении клеточных
мембран, транспорте жиров в организме,
способствуют лучшему усвоению жиров,
препятствуют ожирению печени. Суточная
потребность в фосфолипидах составляет
5 – 10 г.
При усвоении 1
грамма липида выделяется 9 ккал энергии.
При избыточном потреблении жиров
возникает опасность ожирения организма.
Растительные
жиры являются источником поступления
жирорастворимых витаминов Е и β- каротина,
животные жиры – источник жирорастворимых
витаминов А, D.
Тема 5 пищевые кислоты
Продукты питания
содержат различные органические кислоты,
которые объединяют в группу пищевых
кислот. Пищевые кислоты накапливаются
в растительном сырье в результате
биохимических превращений на стадии
развития растения, также кислоты могут
накапливаться вследствие биохимических
изменений в ходе технологического
процесса приготовления продуктов
питания (спиртовое брожение, молочнокислое
брожение). Пищевые кислоты могут быть
внесены в пищевую систему в ходе
технологического процесса для
регулирования рН, придания определенного
вкуса (напитки), для формирования
определенной консистенции (молочные
продукты, кондитерские изделия).
Вносимые пищевые
кислоты в процессе производства продуктов
отнесены к группе пищевых добавок. Их
использование не лимитируется в
гигиеническом отношении, а регламентируется
технологическими инструкциями на
конкретные пищевые продукты. Повышенной
токсичностью обладает фумаровая кислота,
для которой установлен уровень ДСД
допустимой суточной дозы – 6 мг/кг массы
тела человека.
Уксусная
кислота
используется в виде эссенций 70 — 80 %
концентрации и в виде столового уксуса
9 % концентрации. Применяются также соли
уксусной кислоты – ацетаты. Основная
область применения уксусной кислоты –
приготовление овощных консервов.
Молочная
кислота
используется в виде 40 % раствора и
концентрата 70 % раствора. Соли молочной
кислоты называются лактатами. Молочная
кислота применяется в производстве
пива (подкисление затора), безалкогольныз
напитков, кондитерских изделий,
кисломолочных продуктов.
Лимонная
кислота
используется в виде кристаллов белого
цвета, полученных биохимическим синтезом
из плесневого гриба Aspergillus
niger.
Соли лимонной кислоты называются
цитраты. Лимонная кислота имеет мягкий
вкус, меньше раздражает слизистую
оболочку желудочно-кишечного тракта.
В высоких концентрациях лимонная кислота
содержаться в цитрусовых плодах.
Применяется в производстве напитков,
соков, кондитерских изделий, рыбных
консервов.
Яблочная
кислота
используется в виде кристаллов белого
или желтоватого цвета. Соли яблочной
кислоты называются малаты. Яблочная
кислота имеет мягкий вкус, не раздражает
слизистую оболочку желудочно-кишечного
тракта. В высоких концентрациях яблочная
кислота содержаться во фруктах.
Применяется в производстве напитков,
кондитерских изделий.
Винная
кислота
используется в виде кристаллов белого
или желтоватого цвета. Получают при
переработке отходов виноделия. Соли
винной кислоты называются тартраты.
Винная кислота имеет мягкий вкус, меньше
раздражает слизистую оболочку
желудочно-кишечного тракта. Содержится
в винограде. Применяется в производстве
напитков, кондитерских изделий.
Реже
в производстве продуктов питания
используются кислоты: адипиновая,
янтарная, фумаровая.
Фосфорная
кислота
является представителем минеральных
кислот, однако она широко представлена
в в пищевом сырье и продуктах питания,
особенно распространены соли фосфорной
кислоты – фосфаты. Фосфорная кислота
входит в состав сложных органических
соединений: фосфолипиды, нуклеиновые
кислоты, АТФ (аденозинтрифосфат). В
высоких концентрациях фосфаты содержаться
в молочных, мясных продуктах, в орехах.
Применяется в производстве напитков,
кондитерских изделий.
Пищевые
продукты содержат различные аминокислоты: аланин,
валин, серин, лизин, метионин и др.,
входящие в состав белков. Продукты
питания содержат различные липиды, в
состав которых входят жирные
кислоты:
пальмитиновая, стеариновая, олеиновая,
линолевая, линолековая и другие. Ароматическая
кислота –
бензойная кислота является природным
консервантом, она содержится в некоторых
ягодах.
Пищевая ценность липидов — Студопедия
Пищевые жиры и масла являются обязательным компонентом пищи, источником энергетического и пластического материала для человека, поставщиком необходимых веществ, таких как: ненасыщенные жирные кислоты, фосфолипиды, жирорастворимые витамины, стерины. Рекомендуемое содержание жиров в рационе человека по калорийности составляет 30 – 33 % или 90 – 107 г в сутки. Среднем считается норма в 102 г в сутки. В питании имеет значение не только количество, но и химический состав жиров. Линолевая и линоленовые кислоты не синтезируются в организме человека, арахидоновая кислота синтезируется из линолевой кислоты при участии витамина В6. Поэтому они получили название незаменимых или эсенциальных жирных кислот. В последние годы часто употребляется термин «полиненасыщенные жирные кислоты семейства омега – 3», в эту группу входят ά – линоленовая, эйкозапентаеновая, докозагексаеновая кислоты, содержащие несколько кратных связей и «полиненасыщенные жирные кислоты семейства омега – 6», в эту группу вхадит арахидоновая кислота.
Ненасыщенные жирные кислоты участвуют в расщеплении липопротеидов, холестерина, предотвращают образование тромбов, снижают воспалительные процессы.
Липиды оказывают влияние на обмен веществ в клетках, входят в состав клеточных мембран, влияют на кровяное давление, выводят из организма холестерин, при этом повышается эластичность стенок кровеносных сосудов. Повышенной биологической активностью обладают арахидоновая и линолевые кислоты. Среди продуктов питания наиболее богаты полиненасыщенными жирными кислотами растительные масла. Арахидоновая кислота содержится в яйцах, субпродуктах. Сбалансированный состав ежедневного рациона человека должен содержать 10 – 20 % полиненасыщенных жирных кислот, 50 – 60 % мононенасыщенных жирных кислот, 30 % насыщенных жирных кислот. \это обеспечивается при использовании в рационе одной трети растительных и двух третей животных жиров.
Фосфолипиды участвуют в построении клеточных мембран, транспорте жиров в организме, способствуют лучшему усвоению жиров, препятствуют ожирению печени. Суточная потребность в фосфолипидах составляет 5 – 10 г.
При усвоении 1 грамма липида выделяется 9 ккал энергии. При избыточном потреблении жиров возникает опасность ожирения организма.
Растительные жиры являются источником поступления жирорастворимых витаминов Е и β- каротина, животные жиры – источник жирорастворимых витаминов А, D.
2 Пищевая ценность отдельных групп липидов
1.2 Пищевая ценность отдельных групп липидов. Нормы их потребления
Наиболее важная и распространенная группа простых нейтральных липидов – ацилглицеринов. Ацилглицерины – (или глицериды) – это сложные эфиры глицерина и высших карбоновых кислот. Они составляют основную массу липидов (иногда до 95%) и, по существу, именно их называют жирами или маслами. В состав жиров диацилглирерины и моноацилглицерины.
Триацилглицерины (ТАГ), молекулы которых содержат одинаковые остатки жирных кислот, называются простыми, в противном случае — смешанными. Природные жиры и масла содержат, главным образом, смешанные триацилглицерины.
Пищевые жиры относятся к классу липидов, представляющих собой группу соединений животного, растительного или микробного происхождения. Они практически нерастворимы в воде и хорошо растворимы в неполярных органических растворителях. Жиры, добываемые из растительного сырья, называют растительными жирными маслами, а жиры наземных животными жирами. Особую группу составляют жиры морских млекопитающих и рыб.
Чистые ацилглицерины – бесцветные вещества без вкуса и запаха. Окраска, запах и вкус природных жиров определяются наличием в них специфических примесей, характерных для каждого вида жира. Температуры плавления и застывания ацилглицеринов не совпадают, что обусловлено наличием нескольких кристаллических модификаций.
Важнейшая составная часть жиров – жирные кислоты, насыщенные и ненасыщенные (табл. 5.1.).
Таблица 5.1. Основные карбоновые кислоты, входящие в состав природных масел и жиров
Кислота
Рекомендуемые файлы
Формула
Условное обозначение (символ)
Насыщенные кислоты
Лауриновая
Миристиновая
Пальмитиновая
Стеариновая
Арахиновая
СН3-(СН2)10-СООН
СН3-(СН2)12-СООН
СН3-(СН2)14-СООН
СН3-(СН2)16-СООН
СН3-(СН2)18-СООН
С12
С14
С16
С18
С20
Ненасыщенные кислоты
Олеиновая
Эруковая
Линолевая
Линоленовая
Арахидоновая
СН3-(СН2)7-СН=СН-(СН2)7-СООН
СН3-(СН2)7-СН=СН-(СН2)11-СООН
СН3-(СН2)4-СН=СН-СН2-СН=СН-(СН2)7-СООН
СН3-(СН2-СН=СН)3-(СН2)7-СООН
СН3-(СН2)3-(СН2-СН=СН)4-(СН2)3-СООН
С118-9-цис
С122-9-цис
С218-6-цис, 9-цис
С318-3цис, 6-цис, 9-цис
С420-6-цис, 9-цис, 12-цис, 15-цис
*В символ входят число атомов углерода и количество двойных связей между углеродными атомами в молекуле кислоты, номер первого ненасыщенного атома углерода от метильного атома углерода конфигурация.
Жирные кислоты в основном и определяют свойства жира. Чем больше в жирах полиненасыщенных жирных кислот, тем они более биологически активны. Самые распространенные жирные кислоты – пальмитиновая, олеиновая, линолевая.
Насыщенные жирные кислотыиспользуются в основном как энергетический материал, содержатся в наибольших количествах в животных жирах, что определяет высокую температуру плавления этих жиров и их твердое состояние. Они содержатся в мясе животных и субпродуктах.
Высокое содержание животных жиров в рационе нежелательно, поскольку при избытке насыщенных жирных кислот нарушается обмен липидов, повышается уровень холестерина в крови, увеличивается риск развития атеросклероза, ожирения, желчно-каменной болезни.
Ненасыщенные жирные кислоты подразделяются на мононенасыщенные (содержат одну ненасыщенную водородом связь) и полиненасыщенные (несколко связей). Простые ненасыщенные жирные кислоты содержатся в рыбьем жире (эруковая, гадолеиновая), масле, жире, орехах (олеиновая), а также в молочном жире (пальмитолеиновая). Полиненасыщенные жирные кислоты содержатся в масле семян, рыбьем жире (линолевая, линоленовая, арахидоновая, клупонодоновая). Полиненасыщенные жирные кислоты (ПНЖК): линолевая, линоленовая – относятся к незаменимым формам питания, так как в организме они не синтезируются и потому должны поступать с пищей. Эти кислоты по своим биологическим свойствам относятся к жизненно необходимым веществам и называются «Витамин F».
Линолевая кислота превращается в организме в арахидоновую, а линоленовая – эйкозапентаеновую. Недостаточное поступление с пищей линолевой кислоты вызывает в организме нарушение биосинтеза арахидоновой кислоты.
Арахидоновая кислота предшествует образованию веществ, участвующих в регуляции многих процессов жизнедеятельности тромбоцитов и других элементов, но особенно простагландинов, которым придают большое значение как веществам высочайшей биологической активности. Простагландины обладают гормоноподобным действием, в связи с чем получили название «гормонов тканей», так они синтезируются непосредственно из фосфолипидов мембран. Синтез простагландидов зависит от обеспеченности организма этими кислотами.
ПНЖК, образующиеся из линолевой кислоты (эйкозопентановая и докозагексановая), также постоянно в мембранных липидах, но в значительно меньших количествах, чем арахидоновая кислота. ПНЖК участвуют в образовании липидов, вместе с которыми входят в состав клеточных мембран. Воздействуют на структуру кожи и волос, снижают артериальное давление, способствуют профилактике артрита, понижают уровень холестерина и триглицеридов, уменьшают риск тромбообразования; оказывают положительное воздействие при заболеваниях сердечно-сосудистой системы, кандидозе, экземе, псориазе; требуются для нормального развития и функционирования мозга.
Установленная связь ненасыщенных жирных кислот с обменом холестерина. Они способствуют быстрому преобразованию холестерина в фолиевые кислоты и выведению их из организма, оказывают нормализующее действие на стенки кровеносных сосудов, повышают их эластичность и снижают проницаемость. Выявлена зависимость связи ненасыщенных жирных кислот и обмена витаминов группы В. При их дефиците снижается интенсивность роста и устойчивость к неблагоприятным внешним и внутренним факторам, угнетается репродуктивная функция, недостаточность ненасыщенных жирных кислот оказывает влияние на сократительную способность миокарда, вызывает поражение кожи, способствуют развитию атеросклероза. Прием ПНЖК стимулирует систему иммунологической защиты организма, благотворно влияет на внешний вид кожных покровов, способствует более быстрому лечению воспалительных заболеваний желудка, язвенной болезни желудка и двенадцати перстной кишки, способствует оздоровлению и улучшению функции капилляров, эффективен при лечении сахарного диабета и бронхиальной астмы. Особенно много ПНЖК в растительных маслах.
По современным представлениям, сбалансированным считают следующий жирнокислотный состав триацилглицеролов: полиненасыщенные жирные кислоты – 10%, мононенасыщенные – 60%, насыщенные – 30% суточная потребность человека в линолевой кислоте – 4-10 г, что соответствует 20-30 г растительных масел.
По биохимической классификации линолевая кислота и продукты ее превращения объединяются в семейство ω-6 – по положению первой двойной связи в молекуле жирной кислоты, считая от метильного (первого в цепи) атома углерода. Продукты превращения другой незаменимой жирной кислоты – линоленовой – отличаются от представителей жирных кислот семейства ω-6 тем , что у них первая двойная связь от метильного атома углерода занимает положение 3. Поэтому линоленовая кислота и ее продукты превращения образуют семейство ω-3. Жирные кислоты одного семейства в живых организмах не переходят в другое.
На основании современных представлений о физиологической роли ПНЖК разных семейств возникло самостоятельное направление в современной диетологии. Практическим следствием нового направления явилось признание необходимости нормирования и обеспечения постоянного поступления с пищей ПНЖК семейства ω-3. Рассматривается необходимость обеспечения от 0,2 до 0,8% энергоценности рациона за счет линоленовой кислоты, в то время как линолевая кислота (семейство ω-6) должна составлять 4-8% энергоценности. Следовательно, потребность в линоленовой кислоте оценивается в 1/8-1/10 потребности в линолевой. Установлено, что из всех видов растительных масел только соевое имеет соотношение этих двух кислот, близкое к рекомендуемому.
Липиды морских рыб и беспозвоночных содержат главным образом две кислоты семейства ω-3: эйкозапентаеновую и докозагексаеновую. Такой тип липидов получил название «морского». Применение ПНЖК семейства ω-3 в клинике является эффективным методом профилактики атеросклероза и ишемической болезни сердца (ИБС). У больных, перенесших инфаркт миокарда, увеличение содержания в пище линоленовой жирной кислоты в виде изготовленного из рыбьего жира маргарина в течение 5 лет снизило смертность от ИБС на 50%.
Британский фонд питания определил идеальное соотношение в рационе питания человека между ПНЖК семейства ω-6 и ПНЖК ω-3 в количестве 6:1, тогда как, по другим данным, это соотношение должно составлять 10:1. На этом соотношении основана известная рекомендация существенного увеличения потребления жирной рыбы.
Много ненасыщенных жирных кислот содержится в рыбьем жире, в свежей рыбе, в грецких орехах, семенах тыквы, оливках, в льняном, рапсовом масле, примуле вечерней, миндале.
Содержание арахидоновой кислоты в пищевых продуктах незначительно и составляет, %: в мозгах – 0,5; яйцах – 0,1; свиной печени – 0,3; сердце – 0,2. Организмы морских животных , особенно рыб, таких как атерина каспийская, треска, сайра, беломорская и атлантическая сельдь, путассу, антарктический планктонный рачок, голомянка большая, разные виды акул, характеризуются высоким содержанием полиненасыщенных жирных кислот липидной фракции. Наиболее замечательной чертой морских организмов является наличие в их липидных фракциях весьма высоких количеств ПНЖК с 5 и 6 двойными связями. Содержание докозагексаеновой кислоты в жире акулы сельдевой достигает 30%. В общем, в липидах морских организмов содержание высших полиненасыщенных жирных кислот с 4 связями достигает 10%, с 5-30% и с 6-40%.
Важнейшими представителями сложных липидов являются фосфолипиды. Молекулы фосфолипидов построены из остатков спиртов (глицерина, сфиногозина), жирных кислот, фосфорной кислоты ( Н3РО4), а также содержат азотистое основание (чаще всего холин (НО-СН2-СН2-(СН3)3N)+ОН или этаноламин НО-СН2-СН2-NH2), остатки аминокислот и некоторых других соединений.
Фосфолипиды – основной компонент биомембран клеточных структур, они играют существенную роль в проницаемости клеточных оболочек и внутриклеточном обмене. Наиболее важны из фосфолипидов — фосфатидилхолин, или лецитин, проявляет липотропное действие, препятствуя ожирению печени и лучшему усвоению жиров.
Недостаток фосфатидов в рационе приводит к накоплению жира в печени, к ее ожирению, а за тем и к циррозу. Суточная потребность в фосфатидах здорового взрослого человека – 5-10 г.
Лецитин встречается во всех тканях растительного и животного происхождения в семенах масличных растений количество может достигать 1-1,5%, в некоторых тканях животного организма – 6-10%. Лецитином богаты яичные желтки, икра, мозги, печень. Источником лецитина являются, также нерафинированные растительные масла, в том числе и облепиховые, а также молочные жиры. В жирах сливок и сметаны лецитина больше, чем в сливочном масле. В говяжьем, свином, бараньем жирах лецитина почти нет. источником фосфатидов также могут служить бобовые (соя, горох), семена подсолнечника, орехи, особенно кедровые.
При оценке пищевых жиров наиболее высоко ценятся жиры, содержащие лецитин. Для промышленных целей лецитин и кефалин (фосфатидилэтаноламины) получают из соевых бобов. Они используются при производстве шоколада, маргарина и как антиоксиданты в жирах.
Среди сопутствующих жирам неомыляемых веществ важное место занимают стеарины.
Стеарины – алициклические вещества, входящие в группу стероидов, овычно они представляют собой кристаллические одноатомные спирты (стеролы) или их эфиры (стериды). Различают зоостерины, выделяемые из животных объектов, фитостерины (из ратсений), микостерины, выделяемые из грибов. Стерины имеют в своей основе структуру пергидроциклопентанофенантрена
Наиболее известным стерином является холестерин. Он входит в став животных жиров. У млекопитающих он служит предшественником ряда важнейших активных веществ: гормонов, некоторых витаминов, желчных кислот. Холестерин является предшественником гормонов, относящихся к группе стероидных гормонов, в том числе женских половых гормонов прогестерона, эстрадиола и мужского полового гормона тестостерона.
Содержание холестерина в продуктах питания приведено в табл. 5.2.
Таблица 5.2. Содержание холестерина в продуктах
Продукт
Содержание, мг/100 г
Продукт
Содержание, мг/100 г
Мозги
2300
Масло сливочное
240
Яичный желток
1480
Мясо животных и домашней птицы
70
Цельное яйцо
515
Почки
375
Рыба
55
Икра зернистая
более 300
Сыр
90
Печень
300
Творог жирный и сливки
75
При варке мяса и рыбы теряется до 20% холестерина. Обычный суточный рацион – 500 мг холестерина. Известно, что его высокий уровень в крови является фактором риска возникновения атеросклероза, поэтому при соответствующих заболеваниях рекомендуется ограничить потребление пищевых продуктов с его высоким содержанием. В странах, где потребляют наименьшее количество животных жиров (большинство стран Африки, Индия, Япония), содержание холестерина в крови гораздо ниже, чем в США, Англии, Финляндии. Известно, что уменьшение содержания холестерина в крови на 1% приводит к уменьшению риска развития сердечно-сосудистых заболеваний на 2%. Холестерин необходим для синтеза витамина D, желчных кислот, гормонов половых желез и коры надпочечников, а также регуляции проницаемости мембран клеток.
Из фитостеринов, содержащихся в жире растительных продуктов питания, наиболее активным считается β-ситостерин. Он является антогонистом холестерина, задерживает его всасывание в кишечник. Он в больших количествах содержится в растительных маслах. Особенно его много в соевом масле. Β-ситостерол встречается и в мякоти плодов грейпфрута – как в несвязанной форме, так и в виде глюкозида, в семенах грейпфрута он присутствует лишь в свободной форме. Он служит препятствием для абсорбции холестерина, тем самым предотвращая повышение уровня холестерина в сыворотке. Потребление в пищу продуктов, содержащих фитостерин, снижает уровень холестерина в крови.
Рекомендуемое содержание жиров в рационе человека – 90-100 г в сутки, при этом 1/3 их должны составлять растительные масла, 2/3 – животные. По данным ВОЗ, нижний предел безопасного потребления жиров составляет для взрослых мужчин и женщин 25-30 г/сутки.
Информация в лекции «50 Разбой» поможет Вам.
Недостаток или избыток жиров практически одинаково опасны для организма человека (рис. 5.2.). При низком содержании жира в рационе, особенно у людей с нарушенным обменом веществ, сначала появляется сухость и гнойничковые заболевания кожи, затем наступает выпадение волос и нарушение пищеварения, понижается сопротивляемость организма к инфекциям, нарушается обмен витаминов.
При избыточном потреблении жиров происходит их накопление в крови, печени и других тканях и органах. Кровь становится вязкой, повышается ее свертываемость, что предрасполагает к закупорке кровеносных сосудов, возникает атеросклероз. Избыток жира приводит также к ожирению – одному из распространенных заболеваний во многих развитых странах, где потребление жиров на душу населения увеличивается или высока доля жира в традиционных рационах питания.
Рядом ученых высказывается мнение, что существует прямая связь между раком толстого кишечника и потреблением жирной пищи. Высокое содержание жира в пище приводит к увеличению концентрации желчных кислот, поступающих с желчью в кишечник. Желчные кислоты и некоторые другие составные части желчи, а также продукты распада животных белков оказывают на кишечную стенку либо канцерогенное влияние непосредственно, либо под действием кишечной микрофлоры превращаются в продукты, обладающие канцерогенным эффектом. Аналогично этому при избытке ПНЖК, поступающих за счет растительных масел или рыбьих жиров, образуется много окислительных продуктов их обмена – свободных радикалов, — отравляющих печень и почки, снижающих их иммунитет и также оказывающих канцерогенное действие.гфают важную роль в жизнедеятельности организма. Общее количество жира у здорового человека составляет И) — 20 % от массы тела, в случае ожирения может достигать 50%. Существует несколько классов липидов, значительно отличающихся по структуре и по биологическим функциям. Собственно жиры (триглицериды) представляют собой эфиры высших жирных кислот и глицерина. В организме они служат главным источником энергии и образуют резерв энергетического материала. Среди пищевых веществ жиры обладают наибольшей .энергетической ценностью — при сгорании I г жира образуется 9 ккал, при сгорании белков и углеводов -примерно 4 ккал. Во многих тканях даже нри сбалансированном питании для получения энергии используются почти исключительно жиры, тогда как глюкоза сохраняется для тканей, особо в ней нуждающихся (головной мозг, эритроциты). Жиры выполняют защитные фуикиии.
Вокруг жизненно важных орі анов (почки, половые железы, тимус и др.) образуются жировые капсулы, которые удерживают их в нормальном анатомическом положении, предохраняют от смещений и травм. 11а поверхности кожи жиры образуют водоепталкшающую пленку, которая защищает ткани как от потери влаги, так и от переувлажнения, а также обладает антимикробным действием. Кроме зтчлхз подкожный жир создает термоизоляционный покров тела. Жирьг являются плохими проводниками тепла и предохраняют внутренние органы от переохлаждения. Жировая ткань является местом образованна гормона лептина, оказывающего разностороннее воздействие па организм человека. Сложные липиды — это комплексы липидов с бедками (липопрот«иды)> производными орто-фосфорной кислоты (фосфолипиды или фосфатиды), с сахарами (гликолипиды), с многоатомными спиртами (сфинголипиды и др.) и еще целый ряд соединений.
Сложные липиды выполняют пластические функции — опи наряду с белками служат основными структурными компонентами клеточных мембран. К липидам относятся также соединения, не являющиеся производными жирных кислот, — стероиды. Самым распространенным их представителем является холестерин, оті входит как структурный элемент’ в состав клеточных мембран, а также служит предшественником ряда других, стероидов — желчных кислот, стероидных гормонов (гормоны коры надпочечников, половые гормоны), витамина О. Самыми просты-
ми по структуре липидами являются жирные кислоты, которые служат промежуточными продуктами обмена липидов, а также принимают участие в биологической регуляции функций клеток,
Жирпые кислоты могут быть насыщенными и ненасыщенными. В насыщенных кислотах связи между углеродными атомами предельно насыщены, ненасыщенные жирные кислоты содержат одну и более двойных (ненасыщенных) со язей, по месту которых, можеп присоединяться водород. Жирные кислоты с одной двойной связью называются мононенасыщенными (МНЖК). Самой распространенной мононенасыщенной кислотой в жировой ткани человека является олеиновая, срсди других жирных кислот на ее долю приходится 55%, Олеиновая кислота принимает участие в регуляции холестеринового об иена, ОШ способствует повышению В КрОВИ уровня ЛИГЮНТОреИДПВ высокой плотности, ксгторые транспоргируют холсстерин из тканей в печень для утилизации. Этот холестерин не атерогенный или «хороший”, высокий уровень его в крови является фактором антириска для развития атеросклероза. Жирные кислоты с двумя (л и нолевая), тремя (альфа л и ноленовая), четырьмя (арахидо новая) и более двойными связями называются полиненасышенными (ПНЖК).
Ненасыщенные жирные кислоты подразделяют на классы омега в зависимости от положения двойной связи, ближайшей к метил ь но му или омега-углероду, М о ионе насыщенные олеиновая и пальмит иновая кислоты обозначаются соответственно как омега-9 и омега-7, а моли ненасыщенные жирные кислоты линолевая и альфа-линоленовая — как оме га-6 и ом ста-3, Две полиненасыщенные жирные кислоты — линолевая и альф а-шш о ле новая являются незаменимыми (гэссенциальными) для человека, так как они не синтезируются в организме и должны постоянно посту пать извне, из продуктов литания.
У животных, получавших безжировой рацион, отмечено укорочение продолжительности жизни. Особенно негативно сказывается дефицит незаменимых ПНЖК па растутиций организм, линолевая и ап ь фа-лиг юле новая кислота, необходимы для роста, правильного развития головного мозга, органа зрения, половых желез, почек; кожи. Линолевая и альфа-лииоле новая жирные кислоты являются родоначальницами двух семейств ПНЖК — омега-6 и омега-3, в том числе эйко-заноидов — проста гланд инов, лейкотриенов. громбоксанов, являющихся тканевыми гормонами.
Семейство омега-6 представлено линолевой кислотой, которая при наличии необходимых ферментов превращается в организме в г амма-л и но л с новую (ГЛК). Гамма-диноленовая кислота является предшественником дигомогаммалиноленовой кислоты (ДГЛК), которая является предшественницей первой серии простагландинов и арахидоновой кислоты (АК), предшественницы второй серии простагландинов. Альфа-линоленовая кислота превращается в эйкозо-пентасиовую кислоту (ЭПК), предшественницу третьей серии простат а иди но в, и докезагек-саеновую кислот}’ (ДПС). Метаболизм линолевой (18 атомов углерода, две двойных связи) и ал ьфа-л и ноленовой (18 атомов углерода, три двойных связи) кислот связан с дополнительным денасьицением, т.е. увеличением количества двойных связей и удлинением основной цепи, состоящей из атомов углерода, В результате арах идо нова я кислота имеет 20 атомов углерода и 4 двойных связи, эйкозопентае новая геи слота имеет 20 атомов углерода и 5 двойных связей, доко-зогексаеновая кислота имеет 22 атома углерода и 6 двойных связей. Арахидо новую, эйкозопен-таеновую и докозогексасновую кислоты называют длинноцепочными жирными кислотами. Они являются важными структурными компонентами клеточных мембран всех органов и т каней, но особенно велико их содержание в головном мозге, сетчатке глаза, в половых клетках.
Следует отметить, что метаболизм, омега-6 и омега-3 жирных кислот протекает с участием одних и тех же ферментов, что вызывает протекание конкурирующих реакций между двумя этими семействами. Избыток жирных кислот одного класса может тормозить превращения кислот другого класса, снижая их активность и варьируя биологическое действие. Дпинноцепочныс ПНЖК являются предшественниками эйкозаноидов (простагландинов, тромбоксанов, лен котр ненов), выполняющих функции местных тканевых гормонов, которые регулируют многочисленные, функции, включая тонус кровеносных сосудов, мускулатуры бронхов и матки, степень воспалительной реакции, уровень активности клеток иммунной системы, процессы тромбообрязования и ряд других. Причем направленность действия эйкозаноидов семейства омега-6 и омега -3 прямо противоположная. Простагландины, образующиеся из жирных кислот омега-6, суживают просвет4 кровеносных сосудов и бронхов, усиливают воспаление, усиливают тромбообразование. Простагландины -производные жирных кислот оме!а -3 расширяю) бронхи и кровеносные сосуды, уменьшают воспаление. снижают аггрегацию тромбоцитов и уменьшают образование тромбов в кровеносных, сосудах. Иногда первые называют «плохими» простагландинами. а вторые »хорошими». Однако такая оценка является неверной. Именно на принципе противоположности действия основаны процессы
регуляции тканевого гомеостаза. Полому важно не противопоставлять эти два класса жирных кислот а соблюдать баланс между ними. Поскольку источником поли ненасыщенных жирных кислот омега-6 и омега-3 являются пищевые продукты, необходимо знать содержание и соотношение их в рационе. Выполняя стать значимые функции, в организме человека, жиры являются важной составляющей пищевого рациона. Для подаержания оптимального здоровья необходимо придерживаться общих правил рационального питания и потребления жиров, в частности.
Средняя физиологическая потребность в жирах для здорового человека составляем около 30% общей калорийности, третью часть потребляемых жиров до.тжяы составлять растительные .масла. В некоторых специальных диетах долю растительных жиров увеличивают до 50% и более. Жиры улучшают вкус пищи и вызывают чувство сытости, в процессе обмена веществ они могут образовываться из углеводов и белков, но в полной мере ими не заменяются. Пищевая ценность жиров определяется их жирно кислотным составом, наличием незаменимых факторов питания, степенью усвояемости и удобоваримости. Биологическая активность пищевых жиров определяется содержанием в них незаменимых полиненасыщенных жирных кислот. Поскольку основным источником ПНЖК являются растительные масла, го они и обладают наибольшей биологической активностью. Высока и усвояемость растительных масел, в среднем этот показатель составляет 97-98%. Жирные масла растений представляют собой концентрированный энергетический строительный резерв, сосредоточенный в семемах и других органах растений.
Основная роль запасных жиров в растении — использование их для питания во время прорастания семян и развития зародыша; кроме того* они выполняют важную роль защитных кешеств. по-м01*ющих растению переносить неблагоприятные условия окружающей среды, в частности, низкие температуры. Жиры зимующих семян способствуют сохранению зародыша в условиях холода. У деревьев при переходе в состояние покои запасной крахмал превращаемся в жир, повышающий морозостойкость ствола. Наибольшей тепло гворной способностью обладают ненасыщенные жиры, поэтому растения северных широт содержат их в наибольших катичесгвах. Растительные жиры состоят в основном из триглицеридов — эфиров глицерина и жирных кислот. Около 75% растительных жиров со-стаиляют глицериды всег о’грех кислот — пальмитиновой, олеиновой и линолевой.
Жиры некоторых растений содержат специфические, характерные только для них жирные кислоты. Триглицериды мотут быть однокислотными и разнокислотными (смешанными). Од но кислотные жиры (оливковое масло, касторовое масло) встречаются редко, подавляющее большинство жиров представляет собой смеси разнокислотных триглицеридов. Жирные кислоты в растительных жирах могут быть насыщенными и ненасыщенными. Биологическая ценность растительных масел зависит и от содержания в них сопутствующих веществ — фосфолипидов, восков, стероидов, жирорастворимых витаминов, пигментов, фотохимических соединений, содержащихся в растениях и придающих маслам специфическую направленность действия.
Литература:
1 Белобородов В. В.} Основные процессы производства растительных масел, М., 1966.
2. Белобородов В.В., Зайцев Л.С Руководство по методам исследования, технохими-ческому контролю и учету производства в масло-жировой промышленности, т. 5, Л., 1969.
3. Голдовский А. М. Теоретические основы производства растительных масел. М., 1958.
4. Тютюнников Б. Н,, Химия жиров, М., 1966.
5. Щербаков В. Г., Биохимия и товароведение масличного сырья, 2 изд., М., 1969.
это жиры и масла, в состав которых входят жирные кислоты
Обратно в Состав продуктов
По своей структуре липиды представляют собой сложные эфиры глицерина и жирных кислот. Липиды являются одним из важнейших источников энергии для организма. Так, из 1 г липидов организм способен извлечь 9 ккал энергии.
Однако выполняют эти вещества и пластическую функцию (фактически из них состоят мембраны всех клеток и клеточных органелл), и функцию источника предшественников различных физиологически важных веществ (в первую очередь, женских половых гормонов эстрогенов — вот почему очень худые женщины часто страдают репродуктивными нарушениями; кроме того, из непредельных жирных кислот происходит синтез различных медиаторов), а также играют роль переносчика многих гидрофобных соединений, в первую очередь, жирорастворимых витаминов и таких важных минеральных элементов как кальций и магний.
Твердые пищевые липиды принято называть жирами, жидкие — маслами, однако сейчас слово жир преобладает в русском языке и его используют как синоним слову липиды. Жиры и масла являются обязательным компонентом пищи, и их потребность для организма человека определяется на уровне 70-154 г/в сутки и для мужчин и от 60-102 г/сутки для женщин.
Необходимость в них у человека резко возрастает при больших физических нагрузках, а также при нахождении в тяжелых (холодных) климатических условиях.
Жир входит в состав секрета сальных желез, который не только предохраняет кожу от сухости, но и придает волосам прочность и здоровый блеск. У детей жиры служат главным строительным материалом для развития мозга.
В составе пищевых продуктов выделяют видимые и скрытые жиры и масла.
Видимые жиры представляют собой собственно липиды того или иного происхождения, когда человек осознает, что за продукт находиться перед ним. Это растительные жиры и масла, сливочное масло, маргарин, кулинарный жир, сало и т.д.
Скрытые жиры — это те липиды, которые находятся в значительном количестве в пищевых продуктах, но, как правило, потребитель не предполагает, что этот продукт имеют такую жирность. К таким продуктам можно отнести, прежде всего, различные мясные изделия, такие как колбаса, сосиски — содержание жира в них очень велико, а также хлебобулочные и кондитерские изделия.
Разные липиды различаются между собой по происхождению (растительные масла, животные жиры), составу жирных кислот и их расположению (различают альфа- и бета-положение остатка жирных кислот в глицерине; биологической ценностью обладают жирные кислоты, находящиеся в определенном положении; энергетическая же ценность не зависит от положения).
Растительные и животные жиры в рационе
Растительные жиры содержат много ненасыщенных жирных кислот и не содержат холестерин. Обратите внимание, холестерин — это продукт преимущественно животного происхождения, в растительных жирах он либо отсутствует, либо его содержание настолько незначительно, что никогда не выносится на этикетку, т.е. никакое растительное масло не может быть источником холестерина в принципе. Поэтому указание на бутылке растительного масла «0% холестерина», безусловно, не грешит против истины, но по сути, является лишь ловким рекламным ходом, так как растительное масло конкурентов тоже не содержит холестерина. Животные липиды более ценные в плане энергии, а также содержат большее количество жирорастворимых витаминов.
Жирные кислоты различаются между собой длиной цепи, а также наличием (ненасыщенные жирные кислоты) или отсутствием (насыщенные жирные кислоты) двойных связей и их числом (в случае присутствия более одной двойной связи в жирной кислоте говорят о полиненасыщенных жирных кислотах). Типичными примерами насыщенных жирных кислот являются стеариновая, пальмитиновая, маргариновая, лауриновая и др. кислоты; мононенасыщенных — олеиновая, эруковая, вакценовая и др. кислоты; полиненасыщенных — линолевая, линоленовая, арахидоновая и др. кислоты. Чем больше двойных связей в молекуле, тем выше реакционная способность и, соответственно, биологическая ценность масла.
Как насыщенные, так и ненасыщенные жирные кислоты могут быть источником энергетической ценности, в то время как биологической ценностью обладают только ненасыщенные жирные кислоты. Наш организм способен в незначительных количествах синтезировать насыщенные липиды (животные жиры) из углеводов, но ненасыщенные жиры могут поступать в организм человека только с пищей и поэтому относятся к незаменимым факторам питания. Это указывает на необходимость обязательного присутствия в рационе человека ненасыщенных жирных кислот (особенно незаменимых полиненасыщенных жирных кислот омега-3 и омега-6, которые составляют витамин F). Наиболее богаты полиненасыщенными кислотами такие растительные масла как кукурузное, подсолнечное, соевое, льняное и масло грецкого ореха.
По разным причинам не допускается полное исключение жиров и масел из рациона человека, т.к. с ними в организм попадают жирорастворимые витамины (А, Д, Е, К), а жирные кислоты, помимо энергетической, обладают и биологической ценностью. Такой дисбаланс не в пользу жиров часто можно наблюдать при сознательном отказе от них. Это, конечно, иногда и приводит к желаемому результату (похудение), но при этом достигнутый результат сопровождается рядом негативных эффектов: нарушается деятельность центральной нервной системы, снижается устойчивость организма к инфекциям, сокращается продолжительность жизни и пр.
С другой стороны, чрезмерное употребление жиров и углеводов приводит вначале к избыточной массе тела, а потом и вовсе к ожирению. Связано это с тем, что на физиологическом уровне организм ожидает периода голодания. Этот механизм выработался достаточно давно, когда человек в избытке ел только после удачной охоты, после чего следовал долгий период с употреблением скудной пище в ожидании следующей добычи. Поэтому всю избыточную энергетическую ценность (в основном в виде жира и углеводов) организм переводит в жир, который и запасает в специальных клетках жировой ткани — липоцитах.
Так и происходит до тех пор, пока энергетическая ценность потребленных пищевых продуктов и затраченная организмом энергия не сравняется — процесс запасания прекращается. Чтобы начался расход запасенных жиров, необходимо, чтобы расход энергии организмом превышал энергетическую ценность потребленных пищевых продуктов. Это, как правило, достигается коррекцией диеты человека, либо усилением физических нагрузок. Но исключение жиров и масел по причинам указанным выше полностью не допускается, происходить лишь уменьшение количества потребленного жира, а также его замена на биологически более ценный жир (самый простой пример: замена животного жира растительным, характеризующимся высоким содержанием полиненасыщенных жирных кислот).
Обратно в Состав продуктов
Определение чистого энергетического содержания пищевых липидов, скармливаемых растущим свиньям, с помощью непрямой калориметрии
Abstract
Целью этого эксперимента было определение содержания NE в различных пищевых липидах, скармливаемых растущим свиньям, с помощью непрямой калориметрии. Тридцать шесть выращиваемых (исходный BW: 41,1 ± 3,1 кг) курганов были распределены по 6 рационам на основе полностью рандомизированного плана с 6 повторными поросятами на рацион. Диеты включали базальную диету из кукурузо-соевого шрота и 5 тестируемых диет, каждая из которых содержала 10% пальмового масла, птичьего жира, рыбьего жира, кукурузного масла или льняного масла за счет кукурузы и соевого шрота.В течение каждого периода свиней отдельно помещали в клетки для метаболизма на 14 дней, включая 7 дней для адаптации к корму, клеткам для метаболизма и условиям окружающей среды. На 8 день свиней переводили в дыхательные камеры открытого цикла и кормили 1 из 6 рационов из расчета 2,3 МДж ME / кг BW 0,6 / день. Собирали общий кал и мочу, а также рассчитывали ежедневную выработку тепла ( HP ) с 9 по 13 день. В последний день каждого периода (14 день) свиней голодали и производили тепло натощак ( FHP ). был измерен.Результаты показывают, что FHP свиней в среднем составляла 809 кДж / кг BW 0,6 · день -1 и не зависела от характеристик рациона. Значения DE составляли 35,98, 36,84, 37,11, 38,95 и 38,38 МДж / кг сухого вещества, значения ME составляли 35,79, 36,56, 36,92, 37,73 и 38,11 МДж / кг сухого вещества, а значения NE составляли 32,42, 33,21, 33,77, 34,00 и 34,12 МДж / кг сухого вещества для пальмового масла, жира домашней птицы, рыбьего жира, кукурузного масла и льняного масла соответственно. Основываясь на нашем результате, мы пришли к выводу, что содержание DE диетического липида варьировалось от 91% до 98% от его содержания GE, содержание ME диетического липида составляло приблизительно 99% от его содержания DE, а содержание NE в диетическом липиде было приблизительно 90% его МЭ содержится в растущих свиньях.
Ключевые слова: диетические липиды, растущие свиньи, непрямая калориметрия, чистая энергия
ВВЕДЕНИЕ
Липиды, концентрированный источник энергии, обычно добавляют в рацион свиней, а энергетическая ценность липидов примерно в 2,25 раза выше, чем углеводов ( Jones et al., 1992; Lin et al., 2013). Однако энергетическая ценность варьируется в зависимости от источника липидов из-за разного состава жирных кислот, который может влиять на переваривание, абсорбцию и метаболическое использование пищевых липидов (Mendoza and van Heugten, 2014).Также было высказано предположение, что содержание NE в липидах является более точным параметром, чем содержание DE или ME, для описания их продуктивной ценности (Van Heugten et al., 2015). Тем не менее, Sauvant et al. (2004) предложили единое значение NE для всех источников жира (29,77 МДж / кг сухого вещества). В NRC (2012) значение NE липидов было оценено в 88% от ME на основании исследования van Milgen et al. (2001). Было подчеркнуто, что необходимо более точно определять содержание NE в пищевых липидах (Kerr et al., 2015).
Несколько исследователей определили значение NE липидов, используя метод сравнительного убоя ( CS ) (Galloway and Ewan, 1989; Kil et al., 2011). Значение NE для таллового, соевого масла и отборной белой смазки, о которых сообщалось в их исследованиях, было намного меньше, чем у Sauvant et al. (2004) и NRC (2012). Сравнительный метод убоя, который использовал 179 ккал на кг метаболической массы тела (кг 0,6 ) в качестве NE для поддержания (NEm), возможно, недооценил значение NE липидов, обогащенных ненасыщенными жирными кислотами, поскольку NEm, вероятно, будет выше в условиях загона и в связь с повышенным окислительным стрессом (López Bote et al., 2001; Верстеген, 2001). Измерение антиоксидантных параметров может помочь эффективно оценить энергетическую ценность пищевых липидов (Shurson et al., 2015).
Таким образом, целью этого эксперимента было определение содержания NE в источниках липидов с различным составом жирных кислот при скармливании растущим свиньям с использованием непрямой калориметрии ( IC ).
МАТЕРИАЛЫ И МЕТОДЫ
Комитет по контролю за благополучием животных и экспериментальной этикой животных лаборатории Китайского сельскохозяйственного университета (Пекин, Китай) рассмотрел и одобрил все протоколы, использованные в этом эксперименте.
Оборудование для непрямой калориметрии
Шесть респираторных камер открытого цикла, использованные в этом эксперименте, были ранее описаны Liu et al. (2014) и Ли и др. (2017). Температуру поддерживали на уровне 22 ° C в сытом состоянии и постепенно повышали до 24 ° C во время голодания. Относительная влажность поддерживалась на уровне 70%. Содержание кислорода измеряли с помощью парамагнитного дифференциального анализатора (Oxymat 6E, Siemens, Мюнхен, Германия), тогда как содержание CO 2 и CH 4 измеряли с помощью инфракрасных анализаторов (Ultramat 6E, Siemens).Анализаторы имели диапазон измерения от 19,5% до 21% для O 2 , от 0% до 1% для CO 2 и от 0% до 0,1% для CH 4 с чувствительностью 0,2% в пределах измерения. диапазон. Концентрации газа в каждой камере измерялись с 5-минутными интервалами. Мы использовали эксперимент по сжиганию этанола, чтобы проверить точность камеры при измерении газообмена.
Животные, рационы и план эксперимента
Это исследование проводилось в Исследовательском отделении свиней Феннинг Китайского сельскохозяйственного университета (Хэбэй, Китай).Тридцать шесть растущих курганов (дюрок × ландрас × йоркшир) с начальной массой тела 41,1 ± 3,1 кг были распределены на 1 из 6 рационов в соответствии с полностью рандомизированным планом с 6 повторениями свиней на рацион. Каждый раз использовали 6 свиней и 6 дыхательных камер открытого цикла, что дало 6 повторных периодов. Рыбий жир и жир домашней птицы, использованные в этом эксперименте, были кормового качества (свободные жирные кислоты <5%), в то время как все остальные липиды были пищевого качества (свободные жирные кислоты <0,2%). Профиль жирных кислот источников липидов представлен в.Диеты включали базальную диету из кукурузно-соевого шрота ( CNTR ) и 5 тестовых диет, каждая из которых содержала 10% пальмового масла ( PALM ), жир птицы ( POUF ), рыбий жир ( FISH ), кукурузное масло (). CORN ) или льняное масло ( FLAX ) за счет кукурузного и соевого шрота (). Все источники липидов были предоставлены компанией Zhongda Agricultural Science and Technology Co., Ltd. (провинция Шаньдун, Китай). Синтезированный лизин, метионин, треонин, триптофан и валин были добавлены, чтобы они превышали потребности животных во все периоды (NRC, 2012).
Таблица 1.
Анализируемый химический состав источников липидов (%, в исходном состоянии)
Элемент
Пальмовое масло
Жир птицы
Рыбий жир
Кукурузное масло
Льняное масло
Жирные кислоты (% от общего количества жирных кислот)
C8: 0
0,1
0,1
0,1
0,1
0,1
C12: 0
0.1
0,1
0,2
0,0
0,0
C14: 0
0,9
0,8
7,0
0,0
0,1
C16: 0
46,2
23,0
19,8
12,7
5,2
C16: 1
0,1
2,7
7,3
0,1
0,1
C17: 0
0,1
0,2
1.5
0,1
0,1
C18: 0
5,1
8,1
4,2
1,9
3,7
C18: 1 n-9
37,2
39,6
15,0
31,4
28,9
C18: 2 n-6
9,1
21,9
2,0
51,9
16,3
C18: 3 n-6
0,0
0,1
0,4
0.0
0,0
C18: 3 n-3
0,4
1,6
1,8
0,7
43,1
C20: 0
0,4
0,3
0,9
0,4
0,3
C20: 1
0,1
0,5
3,1
0,3
0,8
C20: 2
0,0
0,0
0,1
0,0
0,0
C20 : 3 н-3
ND 5
0.1
0,2
ND
0,1
C20: 3 n-6
ND
0,1
0,2
ND
0,0
C20: 4 n-6
ND
0,1
1,3
ND
0,0
C20: 5 n-3
ND
0,0
12,4
ND
ND
C21: 0
0,0
0,3
0,3
0.0
0,1
C22: 0
0,1
0,2
0,3
0,0
0,2
C22: 1 n-9
0,0
0,0
0,4
0,0
0,8
C22: 2
ND
0,0
0,1
0,0
0,0
C22: 6 n-3
ND
0,0
18,7
ND
ND
C23: 0
0.0
0,0
0,5
0,0
0,0
C24: 0
0,1
0,1
1,5
0,2
0,1
C24: 1
0,0
0,0
0,7
0,0
0,1
PUFA 1
9,4
23,9
37,1
52,7
59,5
УФА 2
46.9
66,8
63,7
84,5
90,1
SFA 3
53,1
33,2
36,2
15,4
9,8
U: S 4
0,9
2,0
1,8
5,5
9,2
Таблица 2.
Ингредиентный состав и химический анализ экспериментальных рационов (%, в исходном состоянии)
Элемент
Диетическое лечение 1
CNTR
PALM
POUF
FISH
CORN
FLAX
Состав,%
Кукуруза
70.37
63,13
63,13
63,13
63,13
63,13
Соевый шрот
26,00
23,32
23,32
23,32
23,32
23,32
Пальмовое масло
—
10.00
—
—
—
—
Птичий жир
—
—
10.00
—
—
—
Рыбий жир
—
—
3 —
—
10.00
—
—
Кукурузное масло
—
—
—
—
10.00
—
Льняное масло
—
—
—
—
10,00
Дикальцийфосфат
1,20
1,20
1,20
1,20
1,20
1,20
Известняк
0.80
0,80
0,80
0,80
0,80
0,80
Соль
0,35
0,35
0,35
0,35
0,35
0,35
Премикс 2 900
0,50
0,50
0,50
0,50
0,50
Lys-HCl 3
0,38
0,34
0,34
0.34
0,34
0,34
DL-Met
0,16
0,14
0,14
0,14
0,14
0,14
L-Thr
0,14
0,13
0,13
0,13
0,13
0,13
L-Trp
0,02
0,02
0,02
0,02
0,02
0,02
L-Val
0.08
0,07
0,07
0,07
0,07
0,07
Анализируемый состав,%
DM
90,07
90,70
90,31
90,36
90,53
89,65
CP
17,49
15,61
15,06
15,01
15,45
15,02
AEE 4
3,83
12.98
12,97
12,52
12,66
12,93
NDF
15,52
13,93
13,67
13,55
13,60
13,83
ADF
3,994 9009 4,4 9009 9,99
3,89
3,66
3,55
Ясень
4,58
4,52
4,44
4,33
4,44
4,39
В течение каждого периода свиньи содержались индивидуально в ящиках для метаболизма в течение 14 d, который включал 7 дней для адаптации к экспериментальной диете, клеткам метаболизма и условиям окружающей среды.На 8 день свиней переводили в дыхательные камеры открытого цикла и кормили 1 из 6 рационов из расчета 2,3 МДж ME / кг BW 0,6 / день. Собирали общий кал и мочу, а также измеряли суточную выработку тепла ( HP ) с 9 по 13 день. В последний день каждого периода (день 14) свиней голодали и производили тепло натощак ( FHP ). измеряли с 22:00 (14-е сутки) до 06:00 (15-е сутки). Период FHP начинался через 30 часов после последнего приема пищи и у животных, которых содержали в темноте, чтобы минимизировать их физическую активность.
Перед экспериментом свиньям скармливали стандартную кукурузно-соевую муку. Свиньи кормили порциями равного размера два раза в день в 09:00 и 16:00. Вода поступала непрерывно через ниппельную поилку низкого давления. Камеры открывали примерно на 1 час в день для кормления свиней и сбора фекалий и мочи. Потребление O 2 и производство CO 2 и CH 4 за это время не были включены в расчет суточного HP. Концентрация CO 2 в камере увеличивалась при закрытии двери.Расчет HP начинался, когда концентрация CO 2 в камере превышала 2000 ppm (Li et al., 2017). Для каждого периода свиней взвешивали на 1, 9, 13 и 14 день. Среднюю массу тела для каждого периода использовали для расчета HP.
Сбор проб
В течение дня с 9 по 13 отбирали остатки корма и просыпи дважды в день для сушки и взвешивания. Общий кал и мочу собирали в соответствии с методами, описанными Liu et al. (2014). Кал собирали дважды в день в 09:00 и 16:00, когда дверь камеры открывали, и сразу хранили при -20 ° C.Мочу собирали отдельно от каждой свиньи в пластиковые ведра, содержащие 50 мл 6 н. HCl, и просеивали через хлопчатобумажную марлю. Был измерен общий объем мочи, произведенной каждой свиньей, и 5% суточной экскреции с мочой сохранялось при -20 ° C. Мочу также собирали отдельно во время 24-часового голодания для расчета потерь азота с мочой для расчета FHP. В конце эксперимента образцы кала и мочи размораживали и тщательно перемешивали, а суб-образец сохраняли для химического анализа.Образцы фекалий сушили в печи в течение 72 часов при 65 ° C. Перед химическим анализом пробы корма и фекалий измельчали через сито диаметром 1 мм. Утром 14-го дня образцы крови (8 мл) были собраны путем пункции передней полой вены с использованием пробирки Vacutainer, не содержащей антикоагулянтов (Becton Dickinson Vacutainer Systems, Franklin Lakes, NJ). Сыворотку получали после центрифугирования (Biofuge22R; Heraeus, Ханау, Германия) при 3000 × g в течение 15 мин при 4 ° C, а затем хранили при -80 ° C до анализа.
Химический анализ
Все химические анализы проводились в двух экземплярах и повторялись, если дубликаты отличались более чем на 5%.Образцы корма и фекалий анализировали на DM (AOAC, 2007; метод 934.01), золу (AOAC 2007; метод 942.15), CP (AOAC, 2007; метод 990.03) и кислотно-гидролизованный эфирный экстракт ( AEE , AOAC, 2007; Метод 954.02). Концентрации NDF и ADF определяли с использованием фильтровальных мешков и оборудования для анализа волокон (Fiber Analyzer, Ankom Technology, Macedon, NY) после модификации процедуры Van Soest et al. (1991). ГЭ в источниках липидов, диетах, образцах мочи и фекалий определяли с помощью калориметра изоперибол (Parr 6300 Calorimeter, Moline, IL) с бензойной кислотой в качестве стандарта.Профили жирных кислот в источниках липидов определяли с помощью газовой хроматографии (серия 6890, Agilent Technologies, Wilmington, DE) по методикам Sukhija и Palmquist (1988) с небольшими изменениями. Образцы липидов были преобразованы в метиловые эфиры жирных кислот с использованием метанольной HCl. Ундекановую кислоту (C11: 0) использовали в качестве внутреннего стандарта. Аликвоты по 1 мкл вводили в капиллярную колонку (60 м × 250 мкм × 250 нм, DB-23, Agilent) с цианопропилметилсиликоном в качестве стационарной фазы.Температуру термостата колонки запрограммировали с разделением 1:20. Температуры инжектора и детектора поддерживались на уровне 260 и 270 ° C соответственно. Азот был газом-носителем при скорости потока 2 мл / мин. Образцы сыворотки размораживали и тщательно перемешивали непосредственно перед анализом. Антиоксидантные параметры, включая супероксиддисмутазу ( SOD ), глутатионпероксидазу ( GSH-PX ), малоновый альдегид ( MDA ), общую антиоксидантную способность ( T-AOC ), каталазу ( CAT ) и соотношение окисленный глутатион ( GSSH ) до общего глутатиона ( T-GSH ) в сыворотке определяли с использованием наборов для анализа в соответствии с инструкциями Beijing Kangjia Bioengineering Company (Пекин, Китай).
Расчеты
Потребление сухого вещества с 9 по 13 день в каждый период рассчитывали как произведение потребления корма и содержания сухого вещества в рационах. Общее потребление энергии рассчитывалось как произведение содержания ГЭ в рационе и фактического потребления сухого вещества корма в течение 5-дневного периода сбора с 9 по 13 день. Энергетические потери с фекалиями, мочой и метаном измерялись для каждого рациона. рассчитывались DE и ME для 6 диет. В ME учитывались потери энергии как с мочой, так и с метаном.Энергия метана была рассчитана с использованием коэффициента преобразования 39,54 кДж / л (Brouwer, 1965).
С 9 по 13 день каждого периода, концентрации O 2 , CO 2 и CH 4 как в входящем, так и в выходящем воздухе, а также скорости выходящего потока воздуха измерялись с 5-минутными интервалами. Эти данные затем использовались для расчета потребления O 2 и производства CO 2 и CH 4 в течение каждого 5-минутного интервала, и эти значения были усреднены и экстраполированы на 24-часовой период.Затем рассчитывалась общая выработка тепла ( THP ) для каждого дня на основе газообмена и потери азота с мочой согласно Брауэру (1965) с использованием уравнения. [1] :):
THP (кДж) = 16,18 × O2 (L) + 5,02 × CO2 (L) — 2,17 × Ch5 (L) — 5,99 × N (г) в моче
(1)
Тепло натощак продуктивность рассчитывалась с использованием того же уравнения, но 24-часовой FHP был спрогнозирован из 8-часового HP после периода отсутствия корма от 31 до 38 часов (с 22:00 до 06:00 часов) в течение последнего дня каждого периода ( Ли и др., 2018). Для базового производства за тот же промежуток времени, что и для расчета THP, 8-часовое HP было также экстраполировано на 24-часовой период. Дыхательный коэффициент ( RQ ) соответствует соотношению между производством CO 2 и потреблением O 2 .
Окисление углеводов ( OXCHO ) и жира ( OXF ) рассчитывали по методу, описанному Chwalibog et al. (1992).
OXCHO (кДж) = [−2,968 × O 2 + 4,147 × CO 2 — 1.761 × CH 4 — 2,446 × N (г) в моче] × 17,58
(2)
OXF (кДж) = [1,719 × O 2 — 1,719 × CO 2 — 1,719 × CH 4 — 1,963 × N (г) в моче] × 39,76
(3)
Кажущуюся общую усвояемость в тракте (ATTD) питательных веществ в рационах рассчитывали в соответствии с методами Noblet et al. (1994).
Сохранение энергии ( RE ) было рассчитано по формуле. [4]:
RE (МДж / кг DM) = [потребление ME (МДж / день) — THP (МДж / день)] / потребление DM (кг / день)
(4)
Сохранение энергии в виде белка ( RE P ) рассчитывали как удерживание N (г) × 6.25 × 23,86 (кДж / г). Сохранение энергии в виде липидов ( RE L ) рассчитывали как разницу между RE и RE P .
Чистая энергия каждого рациона рассчитывалась согласно Noblet et al. (1994) как:
GEc, DEc, Значения MEc и NEc источников липидов были рассчитаны с использованием обновленных методов Adeola (2001) следующим образом:
E липид = [E липидная диета — E базовая диета /96.84 % × 85,70 % ] / 11,19 %
(6)
, где E базальная диета — это среднее значение GE, DE, ME или NE в основной диете (MJ / кг DM ) и соотношение E базальный рацион / 96,84% — это значение GE, DE, ME или NE ингредиентов, которые обеспечивали энергию в базовом рационе. Процент минералов и витаминов в основной диете составлял 3,16% (в пересчете на сухую массу) и не являлся источником энергии.Таким образом, ингредиенты, обеспечивающие энергию (кукуруза, SBM и АК), составляют 96,84% в базовом рационе на основе сухого вещества. Предполагалось, что значения GE, DE, ME и NE смеси кукурузы, SBM и AA, полученные из основного рациона, применимы к другим диетам. E липидная диета — среднее значение GE, DE, ME или NE в липидной диете (МДж / кг СВ). E липид — это расчетные значения GE, DE, ME и NE ( GEc , DEc , MEc, и NEc ) для каждого образца липидов (МДж / кг DM).85,70% — это процент DM ингредиентов (кроме липидов), которые поставляли энергию (кукуруза, SBM и AA) в липидной диете, и 11,19% — это процент DM в образце липидов в липидной диете. DM ингредиентов измеряли перед приготовлением диет, чтобы рассчитать соотношение DM каждого тестируемого ингредиента в рационе. Отношения DEc / GEc, MEc / Dec и NEc / MEc затем могут быть рассчитаны для каждого источника липидов на основе этих рассчитанных значений GEc, DEc, MEc и NEc и использованы для оценки конечных значений DE, ME и NE в качестве продукта. измеренных GEm и DEc / GEc для DE, измеренных GEm и DEc / GEc и MEc / DEc для ME и измеренных GEm и DEc / GEc, MEc / DEc и NEc / MEc для NE (Li et al., 2017). Все расчеты производились на основе DM. Поскольку существует только одно оценочное значение энергии для каждого источника липидов, статистический анализ не может быть проведен среди значений энергии источников липидов.
Статистический анализ
Данные были проверены на нормальность, и выбросы были обнаружены с использованием процедуры UNIVARIATE SAS (Институт SAS, Кэри, Северная Каролина). Никаких выбросов не выявлено. Затем данные были проанализированы с использованием ANOVA с процедурой MIXED SAS. Отдельную свинью рассматривали как экспериментальную единицу, и статистическая модель включала лечебную диету как единственный фиксированный эффект, а период и камеру как случайные эффекты.Средние значения лечения были рассчитаны с использованием инструкции LSMEANS, а статистические различия между видами лечения были разделены с использованием теста Tukey HSD. Во всех анализах различия считались значимыми, если P <0,05, и считались тенденцией при P <0,10.
РЕЗУЛЬТАТЫ
Все свиньи остались здоровыми и без проблем потребляли свой рацион. Фекалии были успешно собраны у всех свиней. ATTD AEE был выше при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими рацион CNTR ( P <0.01; ). Однако на ATTD DM, CP, OM, NDF и ADF в рационах не влияло содержание липидов у растущих свиней.
Таблица 3.
ATTD нутриентов в экспериментальных диетах
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
Коэффициенты усвояемости,%
DM
87.3
87,7
88,4
88,2
90,2
89,4
0,78
0,13
OM
89,1
89,4
89,9
90,1
91,5
90,7
0,71 900 0,18
GE
87,7 b
88,4 ab
89,3 ab
89,8 ab
90,9 a
90.4 ab
0,74
0,04
CP
86,2
85,0
85,7
86,4
88,5
86,3
1,19
0,46
AEE 2 900 b
78,5 a
81,5 a
82,7 a
84,5 a
84,2 a
1,59
<0.01
NDF
70,3
67,6
69,2
68,4
73,3
70,3
2,40
0,63
ADF
62,7
61,9
65,3
66,9 9,1 900
61,9
3,72
0,73
Содержание сырого протеина в диете CNTR было выше, чем в липидной диете, что приводит к большему ( P <0.01) потребление азота и удержание азота у свиней, получавших диету CNTR, по сравнению с свиньями, получавшими липидную диету. Кроме того, свиньи в текущем эксперименте получали аналогичное потребление ME (2300 кДж / кг BW 0,6 / день), поэтому свиньи, получавшие рацион CNTR, имели большее ( P <0,01) потребление DM по сравнению со свиньями, получавшими липидные диеты (). Энергия, потерянная в виде мочи для 6 диет, колебалась от 1,5% до 2,1% ДЭ. По сравнению с диетой CNTR, процент потери DE диеты через CH 4 имел тенденцию быть ниже ( P = 0.06) для липидных диет. Отношение NE к ME среди 6 диет колебалось от 77,2% до 80,6%.
Таблица 4.
Влияние характеристик рациона на энергетический и азотный баланс растущих свиней (на основе СВ)
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
BW, кг
41.67
40,14
40,50
42,40
40,95
41,04
1,10
0,74
Потребление DM, кг / сут
1,40 a
1,23 b
1,23 b
1,23 b
1,23 b
1,24 b
1,23 b
1,21 b
0,02
<0,01
Азотный баланс, г / сут
Впуск
38.9 a
30,4 b
28,0 b
29,6 b
29,0 b
28,7 b
0,71
<0,01
0,71
<0,01
a
4,6 ab
4,0 ab
4,0 ab
3,4 b
3,9 ab
0,38
0,02
Выход мочи3 a
5,3 ab
4,8 ab
4,4 b
4,8 ab
5,5 ab
0,61
0,04
a Удержание 26
20,6 b
19,2 b
21,2 b
20,8 b
19,4 b
0,90
<0,01
900 Энергия мочи,% DE
2.1
1,8
1,7
1,5
2,1
1,8
0,20
0,45
Энергия метана,% DE
0,7
0,5
0,6
0,4
0,7
0,5
0,07
0,06
ME / DE
97,2
97,7
97,6
97,7
97,1
97,7
0,29
0,19
NE / ME
77.2 b
80,3 a
80,4 a
80,6 a
80,4 a
80,3 a
0,76
<0,01
кг BW 0,6 / d
ME потребление,
2,284
2,357
2,332
2,341
2,355
2,343
21,95
0,22
Общее производство тепла
9
a
1,276 b
1,287 b
1,261 b
1,266 b
1,280 b
12.52
<0,01
Окисление углеводов 2
1,610 a
1,313 b
1,265 b
1,261 b
1,2476 b
1,2476 b
37,13
<0,01
Окисление жира 3
-417 b
-159 a
-95 a
-111 925
———— а
-90 а
58.02
<0,01
Общая выработка тепла 4 скорректирована
1,380 a
1,245 b
1,269 b
1,238 b
1,236 9000 900 1,256 b
12,50
<0,01
RE P 5
417 a
335 b
311 b
333 2 b
333 2 б
311 б
11.60
<0,01
RE L 6
496 b
746 a
731 a
746 a
751 2
0 a
24,67
<0,01
Сохранение энергии
913 b
1,081 a
1,042 a
1079 a
1085 a
1085
25.24
<0,01
Производство тепла натощак
819
808
790
802
814
819
11,18
0,44
Состояние дыхания
1,10 a
1,04 b
1,03 b
1,03 b
1,02 b
1,03 b
0.01
<0,01
Быстрое состояние
0,82
0,83
0,82
0,83
0,82
0,82
0,01
0,77
Значения энергии, МДж / кг
74 900 GE
18,09
20,52
20,52
20,55
20,63
20,68
—
—
DE
15,86 b
18.14 а
18,25 а
18,32 а
18,57 а
18,59 а
0,14
<0,01
0,14
<0,01
ME
9295 a
17,82 a
17,90 a
18,03 a
18,16 a
0,15
<0,01
NE
11.90 b
14,23 a
14,33 a
14,43 a
14,49 a
14,58 a
0,17
0,17
0,17
свиней, получавших 6 рационов в ходе текущего эксперимента, составляло 2284, 2357, 2332, 2341, 2355 и 2343 кДж / кг BW 0,6 / день, соответственно, что аналогично ожидаемым значениям для плана эксперимента.Добавление диетического жира, независимо от источника, снизило средний THP и скорректированный THP по сравнению со свиньями, получавшими диету CNTR ( P <0,01). Средние скорректированные THP для 6 диет составляли 1380, 1245, 1269, 1238, 1236 и 1256 кДж / кг BW 0,6 / день, соответственно. Тем не менее, средний FHP составлял 809 кДж / кг BW 0,6 · сут -1 для всех обработок и не зависел от характеристик диеты. У свиней, получавших основной корм в сытом состоянии, было больше ( P <0.01) OXCHO для энергии, чем липидные диеты. RE и RE L были выше при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими рацион CNTR ( P <0,01). Кроме того, добавление диетического жира, независимо от источника, уменьшало RQ в сытом состоянии по сравнению со свиньями, получавшими диету CNTR ( P <0,01), но на средний RQ в состоянии натощак не влияли характеристики диеты.
Не было различий по каким-либо измеренным параметрам сывороточного антиоксиданта (SOD, GSH-PX, MDA, T-AOC, CAT и соотношение GSSH / T-GSH) между 6 группами лечения ().По 5 источникам диетического жира, протестированным в нашем исследовании, определенное соотношение DE к GE составляло от 91% до 98%, определенное отношение ME к DE составляло в среднем 99%, определенное отношение NE к ME составляло в среднем 90%, а определенные значения NE колебалась от 32,42 до 34,12 МДж / кг DM. В количественном отношении пальмовое масло имело более низкие значения DE, ME и NE по сравнению с жиром домашней птицы, рыбьим жиром, кукурузным маслом и льняным маслом ().
Таблица 5.
Влияние характеристик рациона на антиоксидантные параметры сыворотки у растущих свиней
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
Параметры антиоксиданта 2
SOD, ед. / Мл
69
130,38
122,80
123,60
126,11
127,47
4,98
0,66
GSH-PX, Ед / мл
681,67
686,17
654,93
611,52 900,9 634,14
20,45
0,11
МДА, нмоль / мл
5,18
4,13
5,16
4,66
4,73
4,31
0.49
0,59
T-AOC, Ед / мл
8,81
8,62
8,29
9,71
8,77
9,09
0,43
0,54
CAT, Ед / мл
5,79
4,87
4,82
4,87
5,31
5,10
0,42
0,55
GSSH / T-GSH
0,23
0,22
0,23
0,22
0.21
0,25
0,01
0,73
Таблица 6.
Использование энергии и содержание энергии в источнике диетического жира у растущих свиней (на основе СВ)
Элемент
Источник липидов
Пальмовое масло
Жир птицы
Рыбий жир
Кукурузное масло
Льняное масло
Использование энергии 1 ,%
DEc / GEc
91.0
93,4
94,4
98,1
97,5
MEc / DEc
99,5
99,2
99,5
96,9
99,3
NEc / MEc
90,6
90,8
91,5
90,1
89,5
Энергетическая ценность 2 , МДж / кг DM
GEm
39,54
39,43
39,33
39,69
39.36
DE
35,98
36,84
37,11
38,95
38,38
ME
35,79
36,56
36,92
37,73
38,11
NE2
38,11
NE2
33,21
33,77
34,00
34,12
ОБСУЖДЕНИЕ
Усвояемость питательных веществ, энергия и баланс азота
Улучшение ATTD AEE для 5 липидных диет в результате увеличения количества пищевых жиров согласуется с предыдущие наблюдения на растущих свиньях (Jørgensen and Fernandez, 2000; Li et al., 2018), предполагая, что эндогенные потери липидов, связанные с потреблением DM корма, оказывают более сильное влияние на ATTD AEE при низких уровнях рациона, чем на более высоких уровнях, а добавленные липиды имеют большую усвояемость, чем липиды из основного рациона (Jørgensen et al., 1993; Noblet, Perez, 1993; Kil et al., 2010).
Удержание азота было ниже при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими CNTR, в основном из-за более низкого суточного потребления азота и аминокислот.Базальная диета, используемая в текущем исследовании, имела гораздо более высокое соотношение белок / ME, чем диеты с добавлением источников липидов, и можно предположить, что только базальная диета используется для поддержания и набора белка и жира. Жир, включенный в экспериментальную диету, в основном способствует отложению, в результате чего в нашем исследовании в виде жира сохранялось на 50% больше энергии. Следовательно, наш экспериментальный план должен обеспечивать значение NE, которое близко к истинному использованию жира в полноценном рационе. Никаких эффектов источника липидов на RE P и RE L не наблюдалось среди 5 липидных диет.Предыдущая работа с крысами и цыплятами-бройлерами показала, что рационы, обогащенные полиненасыщенными жирными кислотами ( PUFA, ), имеют меньшее отложение общего жира в организме, чем рационы, содержащие насыщенные жирные кислоты (Shimomura et al., 1990; Sanz et al., 2000). Их результаты показали, что крысы и цыплята-бройлеры, получавшие рацион, богатый ПНЖК, имели более высокий уровень катаболизма жиров. Кроме того, эндогенный синтез жира (липогенез) может быть снижен у крыс и цыплят-бройлеров, получавших ненасыщенные жирные кислоты. Однако Чвалибог и Торбек (2000) продемонстрировали, что при нормальных условиях кормления основным источником энергии для растущих свиней являются пищевые углеводы, за которыми следует белок.Углеводы и белок обычно могут обеспечить достаточно энергии, чтобы удовлетворить потребности в поддержании и росте, поэтому диетический жир не должен окисляться, но может задерживаться в организме. Это могло быть одной из возможных причин, объясняющих, почему нам не удалось обнаружить другую картину отложения жира у растущих свиней. В сытом состоянии OXF в этом исследовании был отрицательным, что также указывало на то, что свиньи в сытом состоянии не использовали жир для снабжения энергией, а основным субстратом окисления был углевод (Chwalibog et al., 1992).
Добавление диетического жира, независимо от источника, снизило скорректированный THP по сравнению со свиньями, получавшими диету CNTR. Данные, полученные при выращивании свиней в других исследованиях, также подтверждают наш вывод (Noblet et al., 2001; Li et al., 2018). Это наблюдение связано с более высокой эффективностью использования ME из липидов с преимуществом более низкого приращения тепла по сравнению с эффективностью использования ME из белка, крахмала или волокна (Noblet et al., 1994). Другая причина снижения скорректированного THP может заключаться в том, что HP, связанное с активностью кормления, уменьшается, когда липиды добавляются в рационы, скармливаемые растущим свиньям, что приводит к снижению HP (Li et al., 2018). Однако в нашем исследовании физическая активность не измерялась, и поэтому мы не можем проверить эту гипотезу. Даже без количественной оценки физической активности, FHP в нашем испытании измеряли при минимальной ожидаемой активности (ночью, в темноте, регулировке клетки и т. Д.) И после длительного периода голодания. FHP, измеренный в текущем исследовании, составил в среднем 809 кДж / кг BW 0,6 / день, что близко к значениям FHP, полученным в нашей предыдущей работе с использованием того же метода (Liu et al., 2014; Li et al., 2017), а также близкие к тем, которые были получены группой INRA в соответствии с определенной методологией или использовались в крупномасштабных измерениях NE Noblet et al. (1994). Кроме того, в соответствии с другими литературными данными (Le Bellego et al., 2001; Li et al., 2018), состав диеты не влиял на FHP. Лопес Ботэ и др. (2001) предположили, что ненасыщенные жирные кислоты могут увеличивать окислительный стресс у свиней, что может повышать уровень FHP. Однако в этом исследовании параметры диеты не влияли на параметры сывороточного антиоксиданта.Уменьшение RQ для 5 липидных диет в результате увеличения количества пищевых жиров также согласуется с предыдущими наблюдениями на растущих свиньях (Bruininx et al., 2011; Li et al., 2018). С биохимической точки зрения можно ожидать более высокого RQ в CNTR, потому что RQ для синтеза липидов из глюкозы в значительной степени способствует увеличению общего RQ, тогда как отложение диетического жира в виде липидов тела не влияет на общий RQ (van Milgen , 2002).
Энергетическое содержание дополнительных липидов
В NRC (2012) содержание DE в различных источниках липидов основано на исследованиях Wiseman et al.(1990) и Powles et al. (1993, 1994, 1995) с использованием отношения ненасыщенных жирных кислот к насыщенным жирным кислотам и содержания свободных жирных кислот. Значение ME липидов было оценено как 98% от DE, а значение NE было оценено как 88% от ME на основании исследований van Milgen et al. (2001). Оценки ME и NE в NRC (2012) предполагают, что коэффициент преобразования DE в NE одинаков для всех источников жира. Однако было подчеркнуто, что необходимы дополнительные исследования, чтобы определить, верны ли эти отношения и могут ли они широко применяться к различным источникам жира (Kellner and Patience, 2017).По 5 источникам диетического жира, протестированным в нашем исследовании, отношение DE к GE варьировалось от 91% до 98%, в то время как отношение ME к DE (в среднем 99%) и отношение NE к ME (в среднем 90%) были относительно стабильными среди 5 источников липидов. Несмотря на то, что энергетическая эффективность от DE до NE, которую мы определили, больше, чем оценки NRC (2012), наши результаты по отношениям ME / DE и NE / ME полностью согласуются с результатами Noblet et al. (1994), van Milgen et al. (2001) и Ли и др. (2018). Boyd et al. (2015) также пришли к выводу, что энергетический КПД от DE до NE должен быть выше, чем указано в NRC (2012).У других видов, таких как птица, сообщалось о синергетическом эффекте жира с более высоким расчетным значением ME, чем значение GE, когда жир добавлялся в основной рацион (Cao and Adeola, 2016), что можно объяснить лучшим использованием питательных веществ в рационе. базальная диета при наличии жиров. Однако не было найдено литературы, в которой сообщалось бы о перевариваемости жировой энергии выше 100% у растущих свиней.
Точная оценка энергетической ценности пищевых источников жира не только позволяет производителям свинины оценить финансовую ценность дополнительных липидов, но также поддерживает дифференциацию доступных источников жира (Van Heugten et al., 2015; Келлнер и Пейшенс, 2017). Kerr et al. (2015) также подчеркнули необходимость точного определения содержания NE в пищевых липидах, поскольку системы NE могут позволить более точное прогнозирование энергетической ценности пищевых липидов, чем системы DE и ME (Noblet et al., 1994). Несколько исследователей использовали метод сравнительного убоя для определения значения NE липидов (Galloway and Ewan, 1989; Kil et al., 2011), но значения NE, указанные в их исследованиях (17,49, 20,19 и 25,05 МДж / кг сухого вещества, для жир, соевое масло и отборная белая смазка соответственно) были намного меньше, чем оценки Sauvant et al.(2004) и NRC (2012). Однако по 5 источникам диетического жира, протестированным в этом исследовании, средний определенный NE диетического жира составлял 33,50 МДж / кг сухого вещества, а содержание NE в пальмовом масле было численно самым низким (32,42 МДж / кг сухого вещества) в связи с его самым низким показателем. Отношение DE к GE. Это можно объяснить большим количеством насыщенных жирных кислот в пальмовом масле, которые менее растворимы при воздействии солей желчных кислот и могут уменьшать их включение в смешанные мицеллы и замедлять последующее всасывание (Stahly, 1984; Wiseman et al., 1986). Различные составы жирных кислот не имели численных различий в значениях NE для жира птицы, рыбьего жира, кукурузного масла и льняного масла в этом исследовании. Теоретически, когда в рацион добавляют большое количество насыщенных масел, значительная часть пищевых жирных кислот откладывается непосредственно в тканях организма с энергетической эффективностью 90%. Хотя включены большие количества полиненасыщенных масел, некоторые жирные кислоты могут окисляться с выделением энергии в форме АТФ с энергетической эффективностью 66% (Black, 1995; Kil et al., 2011). Однако Ли и др. (2018) предположили, что большая часть пищевых липидов откладывается непосредственно в жировой ткани без каких-либо биохимических превращений после абсорбции, что также подтверждается Bruininx et al. (2011). Эта гипотеза объясняет отсутствие числовой разницы между NE для жира птицы, рыбьего жира, кукурузного масла и льняного масла и относительно высокое отношение NE к ME для всех липидов, которые использовались в этом эксперименте.
Причина основных расхождений между IC и CS при определении содержания NE в липидах в пище заключается в том, что удержание энергии, измеренное с помощью IC, как правило, больше, чем измеренное с помощью CS (Quiniou et al., 1995). В исследованиях с использованием CS свиньи содержатся в более практичных помещениях, а затем могут свободно передвигаться с большим количеством HP, связанных с физической активностью. Кроме того, свиньи в исследованиях с использованием CS могут иногда подниматься ниже своей критической температуры с снова повышенным HP. Следовательно, HP, измеренное с помощью CS, может быть сильно увеличено в связи с увеличением энергии для поддержания и значений FHP (Verstegen, 2001), в то время как значения FHP, измеренные в нашем исследовании с использованием IC, были получены при минимальной активности, при термонейтральности и т. Д.С этой точки зрения результаты с использованием IC должны быть более надежными. Более того, разные стратегии кормления, генетический фон и возраст свиней между нашим исследованием и исследованием Kil et al. (2011) все они могут влиять на содержание, рост и состав тела свиней, что приводит к вариациям в значениях NE липидов (Boisen and Verstegen, 1998). Чтобы подтвердить значение NE пищевых липидов, Boyd et al. (2015) использовали анализ роста для определения NE выбора белой смазки и предположили, что содержание NE в жирах составляет не менее 32.55 МДж / кг для растущих свиней, что убедительно подтверждает наш текущий результат. Очевидно, что необходимы дальнейшие исследования, чтобы определить НЭ липидов в широком диапазоне разнообразных источников пищевых жиров.
ВЫВОДЫ
Значения NE составили 32,42, 33,21, 33,77, 34,00 и 34,12 МДж / кг сухого вещества для пальмового масла, жира птицы, рыбьего жира, кукурузного масла и льняного масла соответственно. Основываясь на наших результатах, отношения ME / DE (приблизительно 99%) и NE / ME (приблизительно 90%) относительно постоянны для всех источников жира, которые мы определили, в то время как соотношение DE / GE более вариабельно среди различных источников жира у растущих свиней. .Для лучшего использования разнообразных источников жира в свиноводстве необходимо больше работать над усвояемостью жира.
Определение содержания чистой энергии в пищевых липидах, скармливаемых растущим свиньям, с помощью непрямой калориметрии
Abstract
Целью этого эксперимента было определение содержания NE в различных пищевых липидах, скармливаемых растущим свиньям, с помощью непрямой калориметрии. Тридцать шесть выращиваемых (исходный BW: 41,1 ± 3,1 кг) курганов были распределены по 6 рационам на основе полностью рандомизированного плана с 6 повторными поросятами на рацион.Диеты включали базальную диету из кукурузо-соевого шрота и 5 тестируемых диет, каждая из которых содержала 10% пальмового масла, птичьего жира, рыбьего жира, кукурузного масла или льняного масла за счет кукурузы и соевого шрота. В течение каждого периода свиней отдельно помещали в клетки для метаболизма на 14 дней, включая 7 дней для адаптации к корму, клеткам для метаболизма и условиям окружающей среды. На 8 день свиней переводили в дыхательные камеры открытого цикла и кормили 1 из 6 рационов из расчета 2,3 МДж ME / кг BW 0,6 / день. Был собран общий кал и моча, а также рассчитана ежедневная выработка тепла ( HP ) с 9 по 13 день.В последний день каждого периода (день 14) свиней голодали и измеряли выработку тепла натощак ( FHP ). Результаты показывают, что FHP свиней в среднем составляла 809 кДж / кг BW 0,6 · день -1 и не зависела от характеристик рациона. Значения DE составляли 35,98, 36,84, 37,11, 38,95 и 38,38 МДж / кг сухого вещества, значения ME составляли 35,79, 36,56, 36,92, 37,73 и 38,11 МДж / кг сухого вещества, а значения NE составляли 32,42, 33,21, 33,77, 34,00 и 34,12 МДж / кг сухого вещества для пальмового масла, жира домашней птицы, рыбьего жира, кукурузного масла и льняного масла соответственно.Основываясь на нашем результате, мы пришли к выводу, что содержание DE диетического липида варьировалось от 91% до 98% от его содержания GE, содержание ME диетического липида составляло приблизительно 99% от его содержания DE, а содержание NE в диетическом липиде было приблизительно 90% его МЭ содержится в растущих свиньях.
Ключевые слова: пищевых липидов, растущие свиньи, косвенная калориметрия, чистая энергия
ВВЕДЕНИЕ
Липиды, концентрированный источник энергии, обычно добавляют в рацион свиней, а энергетическая ценность липидов составляет около 2.В 25 раз больше, чем углеводов (Jones et al., 1992; Lin et al., 2013). Однако энергетическая ценность варьируется в зависимости от источника липидов из-за разного состава жирных кислот, который может влиять на переваривание, абсорбцию и метаболическое использование пищевых липидов (Mendoza and van Heugten, 2014). Также было высказано предположение, что содержание NE в липидах является более точным параметром, чем содержание DE или ME, для описания их продуктивной ценности (Van Heugten et al., 2015). Тем не менее, Sauvant et al. (2004) предложили единое значение NE для всех источников жира (29.77 МДж / кг DM). В NRC (2012) значение NE липидов было оценено в 88% от ME на основании исследования van Milgen et al. (2001). Подчеркивалось, что необходимо более точно определять содержание НЭ в пищевых липидах (Kerr et al., 2015).
Несколько исследователей определили значение NE липидов, используя метод сравнительного убоя ( CS ) (Galloway and Ewan, 1989; Kil et al., 2011). Значение NE для таллового, соевого масла и отборной белой смазки, о которых сообщалось в их исследованиях, было намного меньше, чем у Sauvant et al.(2004) и NRC (2012). Сравнительный метод убоя, который использовал 179 ккал на кг метаболической массы тела (кг 0,6 ) в качестве NE для поддержания (NEm), возможно, занижал значение NE липидов, обогащенных ненасыщенными жирными кислотами, поскольку NEm, вероятно, будет выше в условиях загона и в связь с повышенным окислительным стрессом (López Bote et al., 2001; Verstegen, 2001). Измерение антиоксидантных параметров может помочь эффективно оценить энергетическую ценность пищевых липидов (Shurson et al., 2015).
Таким образом, целью этого эксперимента было определение содержания NE в источниках липидов с различным составом жирных кислот при скармливании растущим свиньям с использованием непрямой калориметрии ( IC ).
МАТЕРИАЛЫ И МЕТОДЫ
Комитет по контролю за благополучием животных и экспериментальной этикой животных лаборатории Китайского сельскохозяйственного университета (Пекин, Китай) рассмотрел и одобрил все протоколы, использованные в этом эксперименте.
Оборудование для непрямой калориметрии
Шесть респираторных камер открытого цикла, использованные в этом эксперименте, были ранее описаны Liu et al.(2014) и Ли и др. (2017). Температуру поддерживали на уровне 22 ° C в сытом состоянии и постепенно повышали до 24 ° C во время голодания. Относительная влажность поддерживалась на уровне 70%. Содержание кислорода измеряли с помощью парамагнитного дифференциального анализатора (Oxymat 6E, Siemens, Мюнхен, Германия), тогда как содержание CO 2 и CH 4 измеряли с помощью инфракрасных анализаторов (Ultramat 6E, Siemens). Анализаторы имели диапазон измерений от 19,5% до 21% для O 2 , от 0% до 1% для CO 2 и от 0% до 0.1% для CH 4 с чувствительностью 0,2% в пределах диапазона измерения. Концентрации газа в каждой камере измерялись с 5-минутными интервалами. Мы использовали эксперимент по сжиганию этанола, чтобы проверить точность камеры при измерении газообмена.
Животные, рационы и план эксперимента
Это исследование проводилось в Исследовательском отделении свиней Феннинг Китайского сельскохозяйственного университета (Хэбэй, Китай). Тридцать шесть растущих курганов (дюрок × ландрас × йоркшир) с начальной BW 41.1 ± 3,1 кг были выделены на 1 из 6 рационов в соответствии с полностью рандомизированным планом с 6 повторениями свиней на рацион. Каждый раз использовали 6 свиней и 6 дыхательных камер открытого цикла, что дало 6 повторных периодов. Рыбий жир и жир домашней птицы, использованные в этом эксперименте, были кормового качества (свободные жирные кислоты <5%), в то время как все остальные липиды были пищевого качества (свободные жирные кислоты <0,2%). Профиль жирных кислот источников липидов представлен в. Диеты включали базальную диету из кукурузно-соевого шрота ( CNTR ) и 5 тестовых диет, каждая из которых содержала 10% пальмового масла ( PALM ), жир птицы ( POUF ), рыбий жир ( FISH ), кукурузное масло (). CORN ) или льняное масло ( FLAX ) за счет кукурузного и соевого шрота ().Все источники липидов были предоставлены компанией Zhongda Agricultural Science and Technology Co., Ltd. (провинция Шаньдун, Китай). Синтезированный лизин, метионин, треонин, триптофан и валин были добавлены, чтобы они превышали потребности животных во все периоды (NRC, 2012).
Таблица 1.
Анализируемый химический состав источников липидов (%, в исходном состоянии)
Элемент
Пальмовое масло
Жир птицы
Рыбий жир
Кукурузное масло
Льняное масло
Жирные кислоты (% от общего количества жирных кислот)
C8: 0
0.1
0,1
0,1
0,1
0,1
C12: 0
0,1
0,1
0,2
0,0
0,0
C14: 0
0,9
0,8
7,0
0,0
0,1
C16: 0
46,2
23,0
19,8
12,7
5,2
C16: 1
0,1
2,7
7.3
0,1
0,1
C17: 0
0,1
0,2
1,5
0,1
0,1
C18: 0
5,1
8,1
4,2
1,9
3,7
C18: 1 n-9
37,2
39,6
15,0
31,4
28,9
C18: 2 n-6
9,1
21,9
2,0
51.9
16,3
C18: 3 n-6
0,0
0,1
0,4
0,0
0,0
C18: 3 n-3
0,4
1,6
1,8
0,7
43,1
C20: 0
0,4
0,3
0,9
0,4
0,3
C20: 1
0,1
0,5
3,1
0,3
0.8
C20: 2
0,0
0,0
0,1
0,0
0,0
C20: 3 n-3
ND 5
0,1
0,2
ND
0,1
C20: 3 n-6
ND
0,1
0,2
ND
0,0
C20: 4 n-6
ND
0,1
1,3
ND
0,0
C20: 5 n-3
ND
0.0
12,4
ND
ND
C21: 0
0,0
0,3
0,3
0,0
0,1
C22: 0
0,1
0,2
0,3
0,0
0,2
C22: 1 n-9
0,0
0,0
0,4
0,0
0,8
C22: 2
ND
0,0
0,1
0.0
0,0
C22: 6 n-3
ND
0,0
18,7
ND
ND
C23: 0
0,0
0,0
0,5
0,0
0,0
C24: 0
0,1
0,1
1,5
0,2
0,1
C24: 1
0,0
0,0
0,7
0,0
0,1
PUFA 1
9.4
23,9
37,1
52,7
59,5
УФА 2
46,9
66,8
63,7
84,5
90,1
SFA 3
53,1
36,2
15,4
9,8
U: S 4
0,9
2,0
1,8
5,5
9,2
Таблица 2.
Ингредиентный состав и химический анализ экспериментальных рационов (%, в исходном состоянии)
Элемент
Диетическое лечение 1
CNTR
PALM
POUF
FISH
CORN
ЛЕН
Состав,%
Кукуруза
70,37
63,13
63,13
63,13
63,13
63.13
Шрот соевый
26,00
23,32
23,32
23,32
23,32
23,32
Пальмовое масло
—
10,00
—
—
—
— 900
Жир птицы
—
—
10,00
—
—
—
Рыбий жир
—
—
—
10.00
—
—
Кукурузное масло
—
—
—
—
10.00
—
Льняное масло
—
—
—
—
10,00
Дикальцийфосфат
1,20
1,20
1,20
1,20
1,20
1,20
Известняк
0.80
0,80
0,80
0,80
0,80
0,80
Соль
0,35
0,35
0,35
0,35
0,35
0,35
Премикс 2 900
0,50
0,50
0,50
0,50
0,50
Lys-HCl 3
0,38
0,34
0,34
0.34
0,34
0,34
DL-Met
0,16
0,14
0,14
0,14
0,14
0,14
L-Thr
0,14
0,13
0,13
0,13
0,13
0,13
L-Trp
0,02
0,02
0,02
0,02
0,02
0,02
L-Val
0.08
0,07
0,07
0,07
0,07
0,07
Анализируемый состав,%
DM
90,07
90,70
90,31
90,36
90,53
89,65
CP
17,49
15,61
15,06
15,01
15,45
15,02
AEE 4
3,83
12.98
12,97
12,52
12,66
12,93
NDF
15,52
13,93
13,67
13,55
13,60
13,83
ADF
3,994 9009 4,4 9009 9,99
3,89
3,66
3,55
Ясень
4,58
4,52
4,44
4,33
4,44
4,39
В течение каждого периода свиньи содержались индивидуально в ящиках для метаболизма в течение 14 d, который включал 7 дней для адаптации к экспериментальной диете, клеткам метаболизма и условиям окружающей среды.На 8 день свиней переводили в дыхательные камеры открытого цикла и кормили 1 из 6 рационов из расчета 2,3 МДж ME / кг BW 0,6 / день. Собирали общий кал и мочу, а также измеряли суточную выработку тепла ( HP ) с 9 по 13 день. В последний день каждого периода (день 14) свиней голодали и производили тепло натощак ( FHP ). измеряли с 22:00 (14-е сутки) до 06:00 (15-е сутки). Период FHP начинался через 30 часов после последнего приема пищи и у животных, которых содержали в темноте, чтобы минимизировать их физическую активность.
Перед экспериментом свиньям скармливали стандартную кукурузно-соевую муку. Свиньи кормили порциями равного размера два раза в день в 09:00 и 16:00. Вода поступала непрерывно через ниппельную поилку низкого давления. Камеры открывали примерно на 1 час в день для кормления свиней и сбора фекалий и мочи. Потребление O 2 и производство CO 2 и CH 4 за это время не были включены в расчет суточного HP. Концентрация CO 2 в камере увеличивалась при закрытии двери.Расчет HP начинался, когда концентрация CO 2 в камере превышала 2000 ppm (Li et al., 2017). Для каждого периода свиней взвешивали на 1, 9, 13 и 14 день. Среднюю массу тела для каждого периода использовали для расчета HP.
Сбор проб
В течение дня с 9 по 13 отбирали остатки корма и просыпи дважды в день для сушки и взвешивания. Общий кал и мочу собирали в соответствии с методами, описанными Liu et al. (2014). Кал собирали дважды в день в 09:00 и 16:00, когда дверь камеры открывали, и сразу хранили при -20 ° C.Мочу собирали отдельно от каждой свиньи в пластиковые ведра, содержащие 50 мл 6 н. HCl, и просеивали через хлопчатобумажную марлю. Был измерен общий объем мочи, произведенной каждой свиньей, и 5% суточной экскреции с мочой сохранялось при -20 ° C. Мочу также собирали отдельно во время 24-часового голодания для расчета потерь азота с мочой для расчета FHP. В конце эксперимента образцы кала и мочи размораживали и тщательно перемешивали, а суб-образец сохраняли для химического анализа.Образцы фекалий сушили в печи в течение 72 часов при 65 ° C. Перед химическим анализом пробы корма и фекалий измельчали через сито диаметром 1 мм. Утром 14-го дня образцы крови (8 мл) были собраны путем пункции передней полой вены с использованием пробирки Vacutainer, не содержащей антикоагулянтов (Becton Dickinson Vacutainer Systems, Franklin Lakes, NJ). Сыворотку получали после центрифугирования (Biofuge22R; Heraeus, Ханау, Германия) при 3000 × g в течение 15 мин при 4 ° C, а затем хранили при -80 ° C до анализа.
Химический анализ
Все химические анализы проводились в двух экземплярах и повторялись, если дубликаты отличались более чем на 5%.Образцы корма и фекалий анализировали на DM (AOAC, 2007; метод 934.01), золу (AOAC 2007; метод 942.15), CP (AOAC, 2007; метод 990.03) и кислотно-гидролизованный эфирный экстракт ( AEE , AOAC, 2007; Метод 954.02). Концентрации NDF и ADF определяли с использованием фильтровальных мешков и оборудования для анализа волокон (Fiber Analyzer, Ankom Technology, Macedon, NY) после модификации процедуры Van Soest et al. (1991). ГЭ в источниках липидов, диетах, образцах мочи и фекалий определяли с помощью калориметра изоперибол (Parr 6300 Calorimeter, Moline, IL) с бензойной кислотой в качестве стандарта.Профили жирных кислот в источниках липидов определяли с помощью газовой хроматографии (серия 6890, Agilent Technologies, Wilmington, DE) по методикам Sukhija и Palmquist (1988) с небольшими изменениями. Образцы липидов были преобразованы в метиловые эфиры жирных кислот с использованием метанольной HCl. Ундекановую кислоту (C11: 0) использовали в качестве внутреннего стандарта. Аликвоты по 1 мкл вводили в капиллярную колонку (60 м × 250 мкм × 250 нм, DB-23, Agilent) с цианопропилметилсиликоном в качестве стационарной фазы.Температуру термостата колонки запрограммировали с разделением 1:20. Температуры инжектора и детектора поддерживались на уровне 260 и 270 ° C соответственно. Азот был газом-носителем при скорости потока 2 мл / мин. Образцы сыворотки размораживали и тщательно перемешивали непосредственно перед анализом. Антиоксидантные параметры, включая супероксиддисмутазу ( SOD ), глутатионпероксидазу ( GSH-PX ), малоновый альдегид ( MDA ), общую антиоксидантную способность ( T-AOC ), каталазу ( CAT ) и соотношение окисленный глутатион ( GSSH ) до общего глутатиона ( T-GSH ) в сыворотке определяли с использованием наборов для анализа в соответствии с инструкциями Beijing Kangjia Bioengineering Company (Пекин, Китай).
Расчеты
Потребление сухого вещества с 9 по 13 день в каждый период рассчитывали как произведение потребления корма и содержания сухого вещества в рационах. Общее потребление энергии рассчитывалось как произведение содержания ГЭ в рационе и фактического потребления сухого вещества корма в течение 5-дневного периода сбора с 9 по 13 день. Энергетические потери с фекалиями, мочой и метаном измерялись для каждого рациона. рассчитывались DE и ME для 6 диет. В ME учитывались потери энергии как с мочой, так и с метаном.Энергия метана была рассчитана с использованием коэффициента преобразования 39,54 кДж / л (Brouwer, 1965).
С 9 по 13 день каждого периода, концентрации O 2 , CO 2 и CH 4 как в входящем, так и в выходящем воздухе, а также скорости выходящего потока воздуха измерялись с 5-минутными интервалами. Эти данные затем использовались для расчета потребления O 2 и производства CO 2 и CH 4 в течение каждого 5-минутного интервала, и эти значения были усреднены и экстраполированы на 24-часовой период.Затем рассчитывалась общая выработка тепла ( THP ) для каждого дня на основе газообмена и потери азота с мочой согласно Брауэру (1965) с использованием уравнения. [1] :):
THP (кДж) = 16,18 × O2 (L) + 5,02 × CO2 (L) — 2,17 × Ch5 (L) — 5,99 × N (г) в моче
(1)
Тепло натощак продуктивность рассчитывалась с использованием того же уравнения, но 24-часовой FHP был спрогнозирован из 8-часового HP после периода отсутствия корма от 31 до 38 часов (с 22:00 до 06:00 часов) в течение последнего дня каждого периода ( Ли и др., 2018). Для базового производства за тот же промежуток времени, что и для расчета THP, 8-часовое HP было также экстраполировано на 24-часовой период. Дыхательный коэффициент ( RQ ) соответствует соотношению между производством CO 2 и потреблением O 2 .
Окисление углеводов ( OXCHO ) и жира ( OXF ) рассчитывали по методу, описанному Chwalibog et al. (1992).
OXCHO (кДж) = [−2,968 × O 2 + 4,147 × CO 2 — 1.761 × CH 4 — 2,446 × N (г) в моче] × 17,58
(2)
OXF (кДж) = [1,719 × O 2 — 1,719 × CO 2 — 1,719 × CH 4 — 1,963 × N (г) в моче] × 39,76
(3)
Кажущуюся общую усвояемость в тракте (ATTD) питательных веществ в рационах рассчитывали в соответствии с методами Noblet et al. (1994).
Сохранение энергии ( RE ) было рассчитано по формуле. [4]:
RE (МДж / кг DM) = [потребление ME (МДж / день) — THP (МДж / день)] / потребление DM (кг / день)
(4)
Сохранение энергии в виде белка ( RE P ) рассчитывали как удерживание N (г) × 6.25 × 23,86 (кДж / г). Сохранение энергии в виде липидов ( RE L ) рассчитывали как разницу между RE и RE P .
Чистая энергия каждого рациона рассчитывалась согласно Noblet et al. (1994) как:
GEc, DEc, Значения MEc и NEc источников липидов были рассчитаны с использованием обновленных методов Adeola (2001) следующим образом:
E липид = [E липидная диета — E базовая диета /96.84 % × 85,70 % ] / 11,19 %
(6)
, где E базальная диета — это среднее значение GE, DE, ME или NE в основной диете (MJ / кг DM ) и соотношение E базальный рацион / 96,84% — это значение GE, DE, ME или NE ингредиентов, которые обеспечивали энергию в базовом рационе. Процент минералов и витаминов в основной диете составлял 3,16% (в пересчете на сухую массу) и не являлся источником энергии.Таким образом, ингредиенты, обеспечивающие энергию (кукуруза, SBM и АК), составляют 96,84% в базовом рационе на основе сухого вещества. Предполагалось, что значения GE, DE, ME и NE смеси кукурузы, SBM и AA, полученные из основного рациона, применимы к другим диетам. E липидная диета — среднее значение GE, DE, ME или NE в липидной диете (МДж / кг СВ). E липид — это расчетные значения GE, DE, ME и NE ( GEc , DEc , MEc, и NEc ) для каждого образца липидов (МДж / кг DM).85,70% — это процент DM ингредиентов (кроме липидов), которые поставляли энергию (кукуруза, SBM и AA) в липидной диете, и 11,19% — это процент DM в образце липидов в липидной диете. DM ингредиентов измеряли перед приготовлением диет, чтобы рассчитать соотношение DM каждого тестируемого ингредиента в рационе. Отношения DEc / GEc, MEc / Dec и NEc / MEc затем могут быть рассчитаны для каждого источника липидов на основе этих рассчитанных значений GEc, DEc, MEc и NEc и использованы для оценки конечных значений DE, ME и NE в качестве продукта. измеренных GEm и DEc / GEc для DE, измеренных GEm и DEc / GEc и MEc / DEc для ME и измеренных GEm и DEc / GEc, MEc / DEc и NEc / MEc для NE (Li et al., 2017). Все расчеты производились на основе DM. Поскольку существует только одно оценочное значение энергии для каждого источника липидов, статистический анализ не может быть проведен среди значений энергии источников липидов.
Статистический анализ
Данные были проверены на нормальность, и выбросы были обнаружены с использованием процедуры UNIVARIATE SAS (Институт SAS, Кэри, Северная Каролина). Никаких выбросов не выявлено. Затем данные были проанализированы с использованием ANOVA с процедурой MIXED SAS. Отдельную свинью рассматривали как экспериментальную единицу, и статистическая модель включала лечебную диету как единственный фиксированный эффект, а период и камеру как случайные эффекты.Средние значения лечения были рассчитаны с использованием инструкции LSMEANS, а статистические различия между видами лечения были разделены с использованием теста Tukey HSD. Во всех анализах различия считались значимыми, если P <0,05, и считались тенденцией при P <0,10.
РЕЗУЛЬТАТЫ
Все свиньи остались здоровыми и без проблем потребляли свой рацион. Фекалии были успешно собраны у всех свиней. ATTD AEE был выше при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими рацион CNTR ( P <0.01; ). Однако на ATTD DM, CP, OM, NDF и ADF в рационах не влияло содержание липидов у растущих свиней.
Таблица 3.
ATTD нутриентов в экспериментальных диетах
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
Коэффициенты усвояемости,%
DM
87.3
87,7
88,4
88,2
90,2
89,4
0,78
0,13
OM
89,1
89,4
89,9
90,1
91,5
90,7
0,71 900 0,18
GE
87,7 b
88,4 ab
89,3 ab
89,8 ab
90,9 a
90.4 ab
0,74
0,04
CP
86,2
85,0
85,7
86,4
88,5
86,3
1,19
0,46
AEE 2 900 b
78,5 a
81,5 a
82,7 a
84,5 a
84,2 a
1,59
<0.01
NDF
70,3
67,6
69,2
68,4
73,3
70,3
2,40
0,63
ADF
62,7
61,9
65,3
66,9 9,1 900
61,9
3,72
0,73
Содержание сырого протеина в диете CNTR было выше, чем в липидной диете, что приводит к большему ( P <0.01) потребление азота и удержание азота у свиней, получавших диету CNTR, по сравнению с свиньями, получавшими липидную диету. Кроме того, свиньи в текущем эксперименте получали аналогичное потребление ME (2300 кДж / кг BW 0,6 / день), поэтому свиньи, получавшие рацион CNTR, имели большее ( P <0,01) потребление DM по сравнению со свиньями, получавшими липидные диеты (). Энергия, потерянная в виде мочи для 6 диет, колебалась от 1,5% до 2,1% ДЭ. По сравнению с диетой CNTR, процент потери DE диеты через CH 4 имел тенденцию быть ниже ( P = 0.06) для липидных диет. Отношение NE к ME среди 6 диет колебалось от 77,2% до 80,6%.
Таблица 4.
Влияние характеристик рациона на энергетический и азотный баланс растущих свиней (на основе СВ)
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
BW, кг
41.67
40,14
40,50
42,40
40,95
41,04
1,10
0,74
Потребление DM, кг / сут
1,40 a
1,23 b
1,23 b
1,23 b
1,23 b
1,24 b
1,23 b
1,21 b
0,02
<0,01
Азотный баланс, г / сут
Впуск
38.9 a
30,4 b
28,0 b
29,6 b
29,0 b
28,7 b
0,71
<0,01
0,71
<0,01
a
4,6 ab
4,0 ab
4,0 ab
3,4 b
3,9 ab
0,38
0,02
Выход мочи3 a
5,3 ab
4,8 ab
4,4 b
4,8 ab
5,5 ab
0,61
0,04
a Удержание 26
20,6 b
19,2 b
21,2 b
20,8 b
19,4 b
0,90
<0,01
900 Энергия мочи,% DE
2.1
1,8
1,7
1,5
2,1
1,8
0,20
0,45
Энергия метана,% DE
0,7
0,5
0,6
0,4
0,7
0,5
0,07
0,06
ME / DE
97,2
97,7
97,6
97,7
97,1
97,7
0,29
0,19
NE / ME
77.2 b
80,3 a
80,4 a
80,6 a
80,4 a
80,3 a
0,76
<0,01
кг BW 0,6 / d
ME потребление,
2,284
2,357
2,332
2,341
2,355
2,343
21,95
0,22
Общее производство тепла
9
a
1,276 b
1,287 b
1,261 b
1,266 b
1,280 b
12.52
<0,01
Окисление углеводов 2
1,610 a
1,313 b
1,265 b
1,261 b
1,2476 b
1,2476 b
37,13
<0,01
Окисление жира 3
-417 b
-159 a
-95 a
-111 925
———— а
-90 а
58.02
<0,01
Общая выработка тепла 4 скорректирована
1,380 a
1,245 b
1,269 b
1,238 b
1,236 9000 900 1,256 b
12,50
<0,01
RE P 5
417 a
335 b
311 b
333 2 b
333 2 б
311 б
11.60
<0,01
RE L 6
496 b
746 a
731 a
746 a
751 2
0 a
24,67
<0,01
Сохранение энергии
913 b
1,081 a
1,042 a
1079 a
1085 a
1085
25.24
<0,01
Производство тепла натощак
819
808
790
802
814
819
11,18
0,44
Состояние дыхания
1,10 a
1,04 b
1,03 b
1,03 b
1,02 b
1,03 b
0.01
<0,01
Быстрое состояние
0,82
0,83
0,82
0,83
0,82
0,82
0,01
0,77
Значения энергии, МДж / кг
74 900 GE
18,09
20,52
20,52
20,55
20,63
20,68
—
—
DE
15,86 b
18.14 а
18,25 а
18,32 а
18,57 а
18,59 а
0,14
<0,01
0,14
<0,01
ME
9295 a
17,82 a
17,90 a
18,03 a
18,16 a
0,15
<0,01
NE
11.90 b
14,23 a
14,33 a
14,43 a
14,49 a
14,58 a
0,17
0,17
0,17
свиней, получавших 6 рационов в ходе текущего эксперимента, составляло 2284, 2357, 2332, 2341, 2355 и 2343 кДж / кг BW 0,6 / день, соответственно, что аналогично ожидаемым значениям для плана эксперимента.Добавление диетического жира, независимо от источника, снизило средний THP и скорректированный THP по сравнению со свиньями, получавшими диету CNTR ( P <0,01). Средние скорректированные THP для 6 диет составляли 1380, 1245, 1269, 1238, 1236 и 1256 кДж / кг BW 0,6 / день, соответственно. Тем не менее, средний FHP составлял 809 кДж / кг BW 0,6 · сут -1 для всех обработок и не зависел от характеристик диеты. У свиней, получавших основной корм в сытом состоянии, было больше ( P <0.01) OXCHO для энергии, чем липидные диеты. RE и RE L были выше при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими рацион CNTR ( P <0,01). Кроме того, добавление диетического жира, независимо от источника, уменьшало RQ в сытом состоянии по сравнению со свиньями, получавшими диету CNTR ( P <0,01), но на средний RQ в состоянии натощак не влияли характеристики диеты.
Не было различий по каким-либо измеренным параметрам сывороточного антиоксиданта (SOD, GSH-PX, MDA, T-AOC, CAT и соотношение GSSH / T-GSH) между 6 группами лечения ().По 5 источникам диетического жира, протестированным в нашем исследовании, определенное соотношение DE к GE составляло от 91% до 98%, определенное отношение ME к DE составляло в среднем 99%, определенное отношение NE к ME составляло в среднем 90%, а определенные значения NE колебалась от 32,42 до 34,12 МДж / кг DM. В количественном отношении пальмовое масло имело более низкие значения DE, ME и NE по сравнению с жиром домашней птицы, рыбьим жиром, кукурузным маслом и льняным маслом ().
Таблица 5.
Влияние характеристик рациона на антиоксидантные параметры сыворотки у растущих свиней
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
Параметры антиоксиданта 2
SOD, ед. / Мл
69
130,38
122,80
123,60
126,11
127,47
4,98
0,66
GSH-PX, Ед / мл
681,67
686,17
654,93
611,52 900,9 634,14
20,45
0,11
МДА, нмоль / мл
5,18
4,13
5,16
4,66
4,73
4,31
0.49
0,59
T-AOC, Ед / мл
8,81
8,62
8,29
9,71
8,77
9,09
0,43
0,54
CAT, Ед / мл
5,79
4,87
4,82
4,87
5,31
5,10
0,42
0,55
GSSH / T-GSH
0,23
0,22
0,23
0,22
0.21
0,25
0,01
0,73
Таблица 6.
Использование энергии и содержание энергии в источнике диетического жира у растущих свиней (на основе СВ)
Элемент
Источник липидов
Пальмовое масло
Жир птицы
Рыбий жир
Кукурузное масло
Льняное масло
Использование энергии 1 ,%
DEc / GEc
91.0
93,4
94,4
98,1
97,5
MEc / DEc
99,5
99,2
99,5
96,9
99,3
NEc / MEc
90,6
90,8
91,5
90,1
89,5
Энергетическая ценность 2 , МДж / кг DM
GEm
39,54
39,43
39,33
39,69
39.36
DE
35,98
36,84
37,11
38,95
38,38
ME
35,79
36,56
36,92
37,73
38,11
NE2
38,11
NE2
33,21
33,77
34,00
34,12
ОБСУЖДЕНИЕ
Усвояемость питательных веществ, энергия и баланс азота
Улучшение ATTD AEE для 5 липидных диет в результате увеличения количества пищевых жиров согласуется с предыдущие наблюдения на растущих свиньях (Jørgensen and Fernandez, 2000; Li et al., 2018), предполагая, что эндогенные потери липидов, связанные с потреблением DM корма, оказывают более сильное влияние на ATTD AEE при низких уровнях рациона, чем на более высоких уровнях, а добавленные липиды имеют большую усвояемость, чем липиды из основного рациона (Jørgensen et al., 1993; Noblet, Perez, 1993; Kil et al., 2010).
Удержание азота было ниже при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими CNTR, в основном из-за более низкого суточного потребления азота и аминокислот.Базальная диета, используемая в текущем исследовании, имела гораздо более высокое соотношение белок / ME, чем диеты с добавлением источников липидов, и можно предположить, что только базальная диета используется для поддержания и набора белка и жира. Жир, включенный в экспериментальную диету, в основном способствует отложению, в результате чего в нашем исследовании в виде жира сохранялось на 50% больше энергии. Следовательно, наш экспериментальный план должен обеспечивать значение NE, которое близко к истинному использованию жира в полноценном рационе. Никаких эффектов источника липидов на RE P и RE L не наблюдалось среди 5 липидных диет.Предыдущая работа с крысами и цыплятами-бройлерами показала, что рационы, обогащенные полиненасыщенными жирными кислотами ( PUFA, ), имеют меньшее отложение общего жира в организме, чем рационы, содержащие насыщенные жирные кислоты (Shimomura et al., 1990; Sanz et al., 2000). Их результаты показали, что крысы и цыплята-бройлеры, получавшие рацион, богатый ПНЖК, имели более высокий уровень катаболизма жиров. Кроме того, эндогенный синтез жира (липогенез) может быть снижен у крыс и цыплят-бройлеров, получавших ненасыщенные жирные кислоты. Однако Чвалибог и Торбек (2000) продемонстрировали, что при нормальных условиях кормления основным источником энергии для растущих свиней являются пищевые углеводы, за которыми следует белок.Углеводы и белок обычно могут обеспечить достаточно энергии, чтобы удовлетворить потребности в поддержании и росте, поэтому диетический жир не должен окисляться, но может задерживаться в организме. Это могло быть одной из возможных причин, объясняющих, почему нам не удалось обнаружить другую картину отложения жира у растущих свиней. В сытом состоянии OXF в этом исследовании был отрицательным, что также указывало на то, что свиньи в сытом состоянии не использовали жир для снабжения энергией, а основным субстратом окисления был углевод (Chwalibog et al., 1992).
Добавление диетического жира, независимо от источника, снизило скорректированный THP по сравнению со свиньями, получавшими диету CNTR. Данные, полученные при выращивании свиней в других исследованиях, также подтверждают наш вывод (Noblet et al., 2001; Li et al., 2018). Это наблюдение связано с более высокой эффективностью использования ME из липидов с преимуществом более низкого приращения тепла по сравнению с эффективностью использования ME из белка, крахмала или волокна (Noblet et al., 1994). Другая причина снижения скорректированного THP может заключаться в том, что HP, связанное с активностью кормления, уменьшается, когда липиды добавляются в рационы, скармливаемые растущим свиньям, что приводит к снижению HP (Li et al., 2018). Однако в нашем исследовании физическая активность не измерялась, и поэтому мы не можем проверить эту гипотезу. Даже без количественной оценки физической активности, FHP в нашем испытании измеряли при минимальной ожидаемой активности (ночью, в темноте, регулировке клетки и т. Д.) И после длительного периода голодания. FHP, измеренный в текущем исследовании, составил в среднем 809 кДж / кг BW 0,6 / день, что близко к значениям FHP, полученным в нашей предыдущей работе с использованием того же метода (Liu et al., 2014; Li et al., 2017), а также близкие к тем, которые были получены группой INRA в соответствии с определенной методологией или использовались в крупномасштабных измерениях NE Noblet et al. (1994). Кроме того, в соответствии с другими литературными данными (Le Bellego et al., 2001; Li et al., 2018), состав диеты не влиял на FHP. Лопес Ботэ и др. (2001) предположили, что ненасыщенные жирные кислоты могут увеличивать окислительный стресс у свиней, что может повышать уровень FHP. Однако в этом исследовании параметры диеты не влияли на параметры сывороточного антиоксиданта.Уменьшение RQ для 5 липидных диет в результате увеличения количества пищевых жиров также согласуется с предыдущими наблюдениями на растущих свиньях (Bruininx et al., 2011; Li et al., 2018). С биохимической точки зрения можно ожидать более высокого RQ в CNTR, потому что RQ для синтеза липидов из глюкозы в значительной степени способствует увеличению общего RQ, тогда как отложение диетического жира в виде липидов тела не влияет на общий RQ (van Milgen , 2002).
Энергетическое содержание дополнительных липидов
В NRC (2012) содержание DE в различных источниках липидов основано на исследованиях Wiseman et al.(1990) и Powles et al. (1993, 1994, 1995) с использованием отношения ненасыщенных жирных кислот к насыщенным жирным кислотам и содержания свободных жирных кислот. Значение ME липидов было оценено как 98% от DE, а значение NE было оценено как 88% от ME на основании исследований van Milgen et al. (2001). Оценки ME и NE в NRC (2012) предполагают, что коэффициент преобразования DE в NE одинаков для всех источников жира. Однако было подчеркнуто, что необходимы дополнительные исследования, чтобы определить, верны ли эти отношения и могут ли они широко применяться к различным источникам жира (Kellner and Patience, 2017).По 5 источникам диетического жира, протестированным в нашем исследовании, отношение DE к GE варьировалось от 91% до 98%, в то время как отношение ME к DE (в среднем 99%) и отношение NE к ME (в среднем 90%) были относительно стабильными среди 5 источников липидов. Несмотря на то, что энергетическая эффективность от DE до NE, которую мы определили, больше, чем оценки NRC (2012), наши результаты по отношениям ME / DE и NE / ME полностью согласуются с результатами Noblet et al. (1994), van Milgen et al. (2001) и Ли и др. (2018). Boyd et al. (2015) также пришли к выводу, что энергетический КПД от DE до NE должен быть выше, чем указано в NRC (2012).У других видов, таких как птица, сообщалось о синергетическом эффекте жира с более высоким расчетным значением ME, чем значение GE, когда жир добавлялся в основной рацион (Cao and Adeola, 2016), что можно объяснить лучшим использованием питательных веществ в рационе. базальная диета при наличии жиров. Однако не было найдено литературы, в которой сообщалось бы о перевариваемости жировой энергии выше 100% у растущих свиней.
Точная оценка энергетической ценности пищевых источников жира не только позволяет производителям свинины оценить финансовую ценность дополнительных липидов, но также поддерживает дифференциацию доступных источников жира (Van Heugten et al., 2015; Келлнер и Пейшенс, 2017). Kerr et al. (2015) также подчеркнули необходимость точного определения содержания NE в пищевых липидах, поскольку системы NE могут позволить более точное прогнозирование энергетической ценности пищевых липидов, чем системы DE и ME (Noblet et al., 1994). Несколько исследователей использовали метод сравнительного убоя для определения значения NE липидов (Galloway and Ewan, 1989; Kil et al., 2011), но значения NE, указанные в их исследованиях (17,49, 20,19 и 25,05 МДж / кг сухого вещества, для жир, соевое масло и отборная белая смазка соответственно) были намного меньше, чем оценки Sauvant et al.(2004) и NRC (2012). Однако по 5 источникам диетического жира, протестированным в этом исследовании, средний определенный NE диетического жира составлял 33,50 МДж / кг сухого вещества, а содержание NE в пальмовом масле было численно самым низким (32,42 МДж / кг сухого вещества) в связи с его самым низким показателем. Отношение DE к GE. Это можно объяснить большим количеством насыщенных жирных кислот в пальмовом масле, которые менее растворимы при воздействии солей желчных кислот и могут уменьшать их включение в смешанные мицеллы и замедлять последующее всасывание (Stahly, 1984; Wiseman et al., 1986). Различные составы жирных кислот не имели численных различий в значениях NE для жира птицы, рыбьего жира, кукурузного масла и льняного масла в этом исследовании. Теоретически, когда в рацион добавляют большое количество насыщенных масел, значительная часть пищевых жирных кислот откладывается непосредственно в тканях организма с энергетической эффективностью 90%. Хотя включены большие количества полиненасыщенных масел, некоторые жирные кислоты могут окисляться с выделением энергии в форме АТФ с энергетической эффективностью 66% (Black, 1995; Kil et al., 2011). Однако Ли и др. (2018) предположили, что большая часть пищевых липидов откладывается непосредственно в жировой ткани без каких-либо биохимических превращений после абсорбции, что также подтверждается Bruininx et al. (2011). Эта гипотеза объясняет отсутствие числовой разницы между NE для жира птицы, рыбьего жира, кукурузного масла и льняного масла и относительно высокое отношение NE к ME для всех липидов, которые использовались в этом эксперименте.
Причина основных расхождений между IC и CS при определении содержания NE в липидах в пище заключается в том, что удержание энергии, измеренное с помощью IC, как правило, больше, чем измеренное с помощью CS (Quiniou et al., 1995). В исследованиях с использованием CS свиньи содержатся в более практичных помещениях, а затем могут свободно передвигаться с большим количеством HP, связанных с физической активностью. Кроме того, свиньи в исследованиях с использованием CS могут иногда подниматься ниже своей критической температуры с снова повышенным HP. Следовательно, HP, измеренное с помощью CS, может быть сильно увеличено в связи с увеличением энергии для поддержания и значений FHP (Verstegen, 2001), в то время как значения FHP, измеренные в нашем исследовании с использованием IC, были получены при минимальной активности, при термонейтральности и т. Д.С этой точки зрения результаты с использованием IC должны быть более надежными. Более того, разные стратегии кормления, генетический фон и возраст свиней между нашим исследованием и исследованием Kil et al. (2011) все они могут влиять на содержание, рост и состав тела свиней, что приводит к вариациям в значениях NE липидов (Boisen and Verstegen, 1998). Чтобы подтвердить значение NE пищевых липидов, Boyd et al. (2015) использовали анализ роста для определения NE выбора белой смазки и предположили, что содержание NE в жирах составляет не менее 32.55 МДж / кг для растущих свиней, что убедительно подтверждает наш текущий результат. Очевидно, что необходимы дальнейшие исследования, чтобы определить НЭ липидов в широком диапазоне разнообразных источников пищевых жиров.
ВЫВОДЫ
Значения NE составили 32,42, 33,21, 33,77, 34,00 и 34,12 МДж / кг сухого вещества для пальмового масла, жира птицы, рыбьего жира, кукурузного масла и льняного масла соответственно. Основываясь на наших результатах, отношения ME / DE (приблизительно 99%) и NE / ME (приблизительно 90%) относительно постоянны для всех источников жира, которые мы определили, в то время как соотношение DE / GE более вариабельно среди различных источников жира у растущих свиней. .Для лучшего использования разнообразных источников жира в свиноводстве необходимо больше работать над усвояемостью жира.
Определение содержания чистой энергии в пищевых липидах, скармливаемых растущим свиньям, с помощью непрямой калориметрии
Abstract
Целью этого эксперимента было определение содержания NE в различных пищевых липидах, скармливаемых растущим свиньям, с помощью непрямой калориметрии. Тридцать шесть выращиваемых (исходный BW: 41,1 ± 3,1 кг) курганов были распределены по 6 рационам на основе полностью рандомизированного плана с 6 повторными поросятами на рацион.Диеты включали базальную диету из кукурузо-соевого шрота и 5 тестируемых диет, каждая из которых содержала 10% пальмового масла, птичьего жира, рыбьего жира, кукурузного масла или льняного масла за счет кукурузы и соевого шрота. В течение каждого периода свиней отдельно помещали в клетки для метаболизма на 14 дней, включая 7 дней для адаптации к корму, клеткам для метаболизма и условиям окружающей среды. На 8 день свиней переводили в дыхательные камеры открытого цикла и кормили 1 из 6 рационов из расчета 2,3 МДж ME / кг BW 0,6 / день. Был собран общий кал и моча, а также рассчитана ежедневная выработка тепла ( HP ) с 9 по 13 день.В последний день каждого периода (день 14) свиней голодали и измеряли выработку тепла натощак ( FHP ). Результаты показывают, что FHP свиней в среднем составляла 809 кДж / кг BW 0,6 · день -1 и не зависела от характеристик рациона. Значения DE составляли 35,98, 36,84, 37,11, 38,95 и 38,38 МДж / кг сухого вещества, значения ME составляли 35,79, 36,56, 36,92, 37,73 и 38,11 МДж / кг сухого вещества, а значения NE составляли 32,42, 33,21, 33,77, 34,00 и 34,12 МДж / кг сухого вещества для пальмового масла, жира домашней птицы, рыбьего жира, кукурузного масла и льняного масла соответственно.Основываясь на нашем результате, мы пришли к выводу, что содержание DE диетического липида варьировалось от 91% до 98% от его содержания GE, содержание ME диетического липида составляло приблизительно 99% от его содержания DE, а содержание NE в диетическом липиде было приблизительно 90% его МЭ содержится в растущих свиньях.
Ключевые слова: пищевых липидов, растущие свиньи, косвенная калориметрия, чистая энергия
ВВЕДЕНИЕ
Липиды, концентрированный источник энергии, обычно добавляют в рацион свиней, а энергетическая ценность липидов составляет около 2.В 25 раз больше, чем углеводов (Jones et al., 1992; Lin et al., 2013). Однако энергетическая ценность варьируется в зависимости от источника липидов из-за разного состава жирных кислот, который может влиять на переваривание, абсорбцию и метаболическое использование пищевых липидов (Mendoza and van Heugten, 2014). Также было высказано предположение, что содержание NE в липидах является более точным параметром, чем содержание DE или ME, для описания их продуктивной ценности (Van Heugten et al., 2015). Тем не менее, Sauvant et al. (2004) предложили единое значение NE для всех источников жира (29.77 МДж / кг DM). В NRC (2012) значение NE липидов было оценено в 88% от ME на основании исследования van Milgen et al. (2001). Подчеркивалось, что необходимо более точно определять содержание НЭ в пищевых липидах (Kerr et al., 2015).
Несколько исследователей определили значение NE липидов, используя метод сравнительного убоя ( CS ) (Galloway and Ewan, 1989; Kil et al., 2011). Значение NE для таллового, соевого масла и отборной белой смазки, о которых сообщалось в их исследованиях, было намного меньше, чем у Sauvant et al.(2004) и NRC (2012). Сравнительный метод убоя, который использовал 179 ккал на кг метаболической массы тела (кг 0,6 ) в качестве NE для поддержания (NEm), возможно, занижал значение NE липидов, обогащенных ненасыщенными жирными кислотами, поскольку NEm, вероятно, будет выше в условиях загона и в связь с повышенным окислительным стрессом (López Bote et al., 2001; Verstegen, 2001). Измерение антиоксидантных параметров может помочь эффективно оценить энергетическую ценность пищевых липидов (Shurson et al., 2015).
Таким образом, целью этого эксперимента было определение содержания NE в источниках липидов с различным составом жирных кислот при скармливании растущим свиньям с использованием непрямой калориметрии ( IC ).
МАТЕРИАЛЫ И МЕТОДЫ
Комитет по контролю за благополучием животных и экспериментальной этикой животных лаборатории Китайского сельскохозяйственного университета (Пекин, Китай) рассмотрел и одобрил все протоколы, использованные в этом эксперименте.
Оборудование для непрямой калориметрии
Шесть респираторных камер открытого цикла, использованные в этом эксперименте, были ранее описаны Liu et al.(2014) и Ли и др. (2017). Температуру поддерживали на уровне 22 ° C в сытом состоянии и постепенно повышали до 24 ° C во время голодания. Относительная влажность поддерживалась на уровне 70%. Содержание кислорода измеряли с помощью парамагнитного дифференциального анализатора (Oxymat 6E, Siemens, Мюнхен, Германия), тогда как содержание CO 2 и CH 4 измеряли с помощью инфракрасных анализаторов (Ultramat 6E, Siemens). Анализаторы имели диапазон измерений от 19,5% до 21% для O 2 , от 0% до 1% для CO 2 и от 0% до 0.1% для CH 4 с чувствительностью 0,2% в пределах диапазона измерения. Концентрации газа в каждой камере измерялись с 5-минутными интервалами. Мы использовали эксперимент по сжиганию этанола, чтобы проверить точность камеры при измерении газообмена.
Животные, рационы и план эксперимента
Это исследование проводилось в Исследовательском отделении свиней Феннинг Китайского сельскохозяйственного университета (Хэбэй, Китай). Тридцать шесть растущих курганов (дюрок × ландрас × йоркшир) с начальной BW 41.1 ± 3,1 кг были выделены на 1 из 6 рационов в соответствии с полностью рандомизированным планом с 6 повторениями свиней на рацион. Каждый раз использовали 6 свиней и 6 дыхательных камер открытого цикла, что дало 6 повторных периодов. Рыбий жир и жир домашней птицы, использованные в этом эксперименте, были кормового качества (свободные жирные кислоты <5%), в то время как все остальные липиды были пищевого качества (свободные жирные кислоты <0,2%). Профиль жирных кислот источников липидов представлен в. Диеты включали базальную диету из кукурузно-соевого шрота ( CNTR ) и 5 тестовых диет, каждая из которых содержала 10% пальмового масла ( PALM ), жир птицы ( POUF ), рыбий жир ( FISH ), кукурузное масло (). CORN ) или льняное масло ( FLAX ) за счет кукурузного и соевого шрота ().Все источники липидов были предоставлены компанией Zhongda Agricultural Science and Technology Co., Ltd. (провинция Шаньдун, Китай). Синтезированный лизин, метионин, треонин, триптофан и валин были добавлены, чтобы они превышали потребности животных во все периоды (NRC, 2012).
Таблица 1.
Анализируемый химический состав источников липидов (%, в исходном состоянии)
Элемент
Пальмовое масло
Жир птицы
Рыбий жир
Кукурузное масло
Льняное масло
Жирные кислоты (% от общего количества жирных кислот)
C8: 0
0.1
0,1
0,1
0,1
0,1
C12: 0
0,1
0,1
0,2
0,0
0,0
C14: 0
0,9
0,8
7,0
0,0
0,1
C16: 0
46,2
23,0
19,8
12,7
5,2
C16: 1
0,1
2,7
7.3
0,1
0,1
C17: 0
0,1
0,2
1,5
0,1
0,1
C18: 0
5,1
8,1
4,2
1,9
3,7
C18: 1 n-9
37,2
39,6
15,0
31,4
28,9
C18: 2 n-6
9,1
21,9
2,0
51.9
16,3
C18: 3 n-6
0,0
0,1
0,4
0,0
0,0
C18: 3 n-3
0,4
1,6
1,8
0,7
43,1
C20: 0
0,4
0,3
0,9
0,4
0,3
C20: 1
0,1
0,5
3,1
0,3
0.8
C20: 2
0,0
0,0
0,1
0,0
0,0
C20: 3 n-3
ND 5
0,1
0,2
ND
0,1
C20: 3 n-6
ND
0,1
0,2
ND
0,0
C20: 4 n-6
ND
0,1
1,3
ND
0,0
C20: 5 n-3
ND
0.0
12,4
ND
ND
C21: 0
0,0
0,3
0,3
0,0
0,1
C22: 0
0,1
0,2
0,3
0,0
0,2
C22: 1 n-9
0,0
0,0
0,4
0,0
0,8
C22: 2
ND
0,0
0,1
0.0
0,0
C22: 6 n-3
ND
0,0
18,7
ND
ND
C23: 0
0,0
0,0
0,5
0,0
0,0
C24: 0
0,1
0,1
1,5
0,2
0,1
C24: 1
0,0
0,0
0,7
0,0
0,1
PUFA 1
9.4
23,9
37,1
52,7
59,5
УФА 2
46,9
66,8
63,7
84,5
90,1
SFA 3
53,1
36,2
15,4
9,8
U: S 4
0,9
2,0
1,8
5,5
9,2
Таблица 2.
Ингредиентный состав и химический анализ экспериментальных рационов (%, в исходном состоянии)
Элемент
Диетическое лечение 1
CNTR
PALM
POUF
FISH
CORN
ЛЕН
Состав,%
Кукуруза
70,37
63,13
63,13
63,13
63,13
63.13
Шрот соевый
26,00
23,32
23,32
23,32
23,32
23,32
Пальмовое масло
—
10,00
—
—
—
— 900
Жир птицы
—
—
10,00
—
—
—
Рыбий жир
—
—
—
10.00
—
—
Кукурузное масло
—
—
—
—
10.00
—
Льняное масло
—
—
—
—
10,00
Дикальцийфосфат
1,20
1,20
1,20
1,20
1,20
1,20
Известняк
0.80
0,80
0,80
0,80
0,80
0,80
Соль
0,35
0,35
0,35
0,35
0,35
0,35
Премикс 2 900
0,50
0,50
0,50
0,50
0,50
Lys-HCl 3
0,38
0,34
0,34
0.34
0,34
0,34
DL-Met
0,16
0,14
0,14
0,14
0,14
0,14
L-Thr
0,14
0,13
0,13
0,13
0,13
0,13
L-Trp
0,02
0,02
0,02
0,02
0,02
0,02
L-Val
0.08
0,07
0,07
0,07
0,07
0,07
Анализируемый состав,%
DM
90,07
90,70
90,31
90,36
90,53
89,65
CP
17,49
15,61
15,06
15,01
15,45
15,02
AEE 4
3,83
12.98
12,97
12,52
12,66
12,93
NDF
15,52
13,93
13,67
13,55
13,60
13,83
ADF
3,994 9009 4,4 9009 9,99
3,89
3,66
3,55
Ясень
4,58
4,52
4,44
4,33
4,44
4,39
В течение каждого периода свиньи содержались индивидуально в ящиках для метаболизма в течение 14 d, который включал 7 дней для адаптации к экспериментальной диете, клеткам метаболизма и условиям окружающей среды.На 8 день свиней переводили в дыхательные камеры открытого цикла и кормили 1 из 6 рационов из расчета 2,3 МДж ME / кг BW 0,6 / день. Собирали общий кал и мочу, а также измеряли суточную выработку тепла ( HP ) с 9 по 13 день. В последний день каждого периода (день 14) свиней голодали и производили тепло натощак ( FHP ). измеряли с 22:00 (14-е сутки) до 06:00 (15-е сутки). Период FHP начинался через 30 часов после последнего приема пищи и у животных, которых содержали в темноте, чтобы минимизировать их физическую активность.
Перед экспериментом свиньям скармливали стандартную кукурузно-соевую муку. Свиньи кормили порциями равного размера два раза в день в 09:00 и 16:00. Вода поступала непрерывно через ниппельную поилку низкого давления. Камеры открывали примерно на 1 час в день для кормления свиней и сбора фекалий и мочи. Потребление O 2 и производство CO 2 и CH 4 за это время не были включены в расчет суточного HP. Концентрация CO 2 в камере увеличивалась при закрытии двери.Расчет HP начинался, когда концентрация CO 2 в камере превышала 2000 ppm (Li et al., 2017). Для каждого периода свиней взвешивали на 1, 9, 13 и 14 день. Среднюю массу тела для каждого периода использовали для расчета HP.
Сбор проб
В течение дня с 9 по 13 отбирали остатки корма и просыпи дважды в день для сушки и взвешивания. Общий кал и мочу собирали в соответствии с методами, описанными Liu et al. (2014). Кал собирали дважды в день в 09:00 и 16:00, когда дверь камеры открывали, и сразу хранили при -20 ° C.Мочу собирали отдельно от каждой свиньи в пластиковые ведра, содержащие 50 мл 6 н. HCl, и просеивали через хлопчатобумажную марлю. Был измерен общий объем мочи, произведенной каждой свиньей, и 5% суточной экскреции с мочой сохранялось при -20 ° C. Мочу также собирали отдельно во время 24-часового голодания для расчета потерь азота с мочой для расчета FHP. В конце эксперимента образцы кала и мочи размораживали и тщательно перемешивали, а суб-образец сохраняли для химического анализа.Образцы фекалий сушили в печи в течение 72 часов при 65 ° C. Перед химическим анализом пробы корма и фекалий измельчали через сито диаметром 1 мм. Утром 14-го дня образцы крови (8 мл) были собраны путем пункции передней полой вены с использованием пробирки Vacutainer, не содержащей антикоагулянтов (Becton Dickinson Vacutainer Systems, Franklin Lakes, NJ). Сыворотку получали после центрифугирования (Biofuge22R; Heraeus, Ханау, Германия) при 3000 × g в течение 15 мин при 4 ° C, а затем хранили при -80 ° C до анализа.
Химический анализ
Все химические анализы проводились в двух экземплярах и повторялись, если дубликаты отличались более чем на 5%.Образцы корма и фекалий анализировали на DM (AOAC, 2007; метод 934.01), золу (AOAC 2007; метод 942.15), CP (AOAC, 2007; метод 990.03) и кислотно-гидролизованный эфирный экстракт ( AEE , AOAC, 2007; Метод 954.02). Концентрации NDF и ADF определяли с использованием фильтровальных мешков и оборудования для анализа волокон (Fiber Analyzer, Ankom Technology, Macedon, NY) после модификации процедуры Van Soest et al. (1991). ГЭ в источниках липидов, диетах, образцах мочи и фекалий определяли с помощью калориметра изоперибол (Parr 6300 Calorimeter, Moline, IL) с бензойной кислотой в качестве стандарта.Профили жирных кислот в источниках липидов определяли с помощью газовой хроматографии (серия 6890, Agilent Technologies, Wilmington, DE) по методикам Sukhija и Palmquist (1988) с небольшими изменениями. Образцы липидов были преобразованы в метиловые эфиры жирных кислот с использованием метанольной HCl. Ундекановую кислоту (C11: 0) использовали в качестве внутреннего стандарта. Аликвоты по 1 мкл вводили в капиллярную колонку (60 м × 250 мкм × 250 нм, DB-23, Agilent) с цианопропилметилсиликоном в качестве стационарной фазы.Температуру термостата колонки запрограммировали с разделением 1:20. Температуры инжектора и детектора поддерживались на уровне 260 и 270 ° C соответственно. Азот был газом-носителем при скорости потока 2 мл / мин. Образцы сыворотки размораживали и тщательно перемешивали непосредственно перед анализом. Антиоксидантные параметры, включая супероксиддисмутазу ( SOD ), глутатионпероксидазу ( GSH-PX ), малоновый альдегид ( MDA ), общую антиоксидантную способность ( T-AOC ), каталазу ( CAT ) и соотношение окисленный глутатион ( GSSH ) до общего глутатиона ( T-GSH ) в сыворотке определяли с использованием наборов для анализа в соответствии с инструкциями Beijing Kangjia Bioengineering Company (Пекин, Китай).
Расчеты
Потребление сухого вещества с 9 по 13 день в каждый период рассчитывали как произведение потребления корма и содержания сухого вещества в рационах. Общее потребление энергии рассчитывалось как произведение содержания ГЭ в рационе и фактического потребления сухого вещества корма в течение 5-дневного периода сбора с 9 по 13 день. Энергетические потери с фекалиями, мочой и метаном измерялись для каждого рациона. рассчитывались DE и ME для 6 диет. В ME учитывались потери энергии как с мочой, так и с метаном.Энергия метана была рассчитана с использованием коэффициента преобразования 39,54 кДж / л (Brouwer, 1965).
С 9 по 13 день каждого периода, концентрации O 2 , CO 2 и CH 4 как в входящем, так и в выходящем воздухе, а также скорости выходящего потока воздуха измерялись с 5-минутными интервалами. Эти данные затем использовались для расчета потребления O 2 и производства CO 2 и CH 4 в течение каждого 5-минутного интервала, и эти значения были усреднены и экстраполированы на 24-часовой период.Затем рассчитывалась общая выработка тепла ( THP ) для каждого дня на основе газообмена и потери азота с мочой согласно Брауэру (1965) с использованием уравнения. [1] :):
THP (кДж) = 16,18 × O2 (L) + 5,02 × CO2 (L) — 2,17 × Ch5 (L) — 5,99 × N (г) в моче
(1)
Тепло натощак продуктивность рассчитывалась с использованием того же уравнения, но 24-часовой FHP был спрогнозирован из 8-часового HP после периода отсутствия корма от 31 до 38 часов (с 22:00 до 06:00 часов) в течение последнего дня каждого периода ( Ли и др., 2018). Для базового производства за тот же промежуток времени, что и для расчета THP, 8-часовое HP было также экстраполировано на 24-часовой период. Дыхательный коэффициент ( RQ ) соответствует соотношению между производством CO 2 и потреблением O 2 .
Окисление углеводов ( OXCHO ) и жира ( OXF ) рассчитывали по методу, описанному Chwalibog et al. (1992).
OXCHO (кДж) = [−2,968 × O 2 + 4,147 × CO 2 — 1.761 × CH 4 — 2,446 × N (г) в моче] × 17,58
(2)
OXF (кДж) = [1,719 × O 2 — 1,719 × CO 2 — 1,719 × CH 4 — 1,963 × N (г) в моче] × 39,76
(3)
Кажущуюся общую усвояемость в тракте (ATTD) питательных веществ в рационах рассчитывали в соответствии с методами Noblet et al. (1994).
Сохранение энергии ( RE ) было рассчитано по формуле. [4]:
RE (МДж / кг DM) = [потребление ME (МДж / день) — THP (МДж / день)] / потребление DM (кг / день)
(4)
Сохранение энергии в виде белка ( RE P ) рассчитывали как удерживание N (г) × 6.25 × 23,86 (кДж / г). Сохранение энергии в виде липидов ( RE L ) рассчитывали как разницу между RE и RE P .
Чистая энергия каждого рациона рассчитывалась согласно Noblet et al. (1994) как:
GEc, DEc, Значения MEc и NEc источников липидов были рассчитаны с использованием обновленных методов Adeola (2001) следующим образом:
E липид = [E липидная диета — E базовая диета /96.84 % × 85,70 % ] / 11,19 %
(6)
, где E базальная диета — это среднее значение GE, DE, ME или NE в основной диете (MJ / кг DM ) и соотношение E базальный рацион / 96,84% — это значение GE, DE, ME или NE ингредиентов, которые обеспечивали энергию в базовом рационе. Процент минералов и витаминов в основной диете составлял 3,16% (в пересчете на сухую массу) и не являлся источником энергии.Таким образом, ингредиенты, обеспечивающие энергию (кукуруза, SBM и АК), составляют 96,84% в базовом рационе на основе сухого вещества. Предполагалось, что значения GE, DE, ME и NE смеси кукурузы, SBM и AA, полученные из основного рациона, применимы к другим диетам. E липидная диета — среднее значение GE, DE, ME или NE в липидной диете (МДж / кг СВ). E липид — это расчетные значения GE, DE, ME и NE ( GEc , DEc , MEc, и NEc ) для каждого образца липидов (МДж / кг DM).85,70% — это процент DM ингредиентов (кроме липидов), которые поставляли энергию (кукуруза, SBM и AA) в липидной диете, и 11,19% — это процент DM в образце липидов в липидной диете. DM ингредиентов измеряли перед приготовлением диет, чтобы рассчитать соотношение DM каждого тестируемого ингредиента в рационе. Отношения DEc / GEc, MEc / Dec и NEc / MEc затем могут быть рассчитаны для каждого источника липидов на основе этих рассчитанных значений GEc, DEc, MEc и NEc и использованы для оценки конечных значений DE, ME и NE в качестве продукта. измеренных GEm и DEc / GEc для DE, измеренных GEm и DEc / GEc и MEc / DEc для ME и измеренных GEm и DEc / GEc, MEc / DEc и NEc / MEc для NE (Li et al., 2017). Все расчеты производились на основе DM. Поскольку существует только одно оценочное значение энергии для каждого источника липидов, статистический анализ не может быть проведен среди значений энергии источников липидов.
Статистический анализ
Данные были проверены на нормальность, и выбросы были обнаружены с использованием процедуры UNIVARIATE SAS (Институт SAS, Кэри, Северная Каролина). Никаких выбросов не выявлено. Затем данные были проанализированы с использованием ANOVA с процедурой MIXED SAS. Отдельную свинью рассматривали как экспериментальную единицу, и статистическая модель включала лечебную диету как единственный фиксированный эффект, а период и камеру как случайные эффекты.Средние значения лечения были рассчитаны с использованием инструкции LSMEANS, а статистические различия между видами лечения были разделены с использованием теста Tukey HSD. Во всех анализах различия считались значимыми, если P <0,05, и считались тенденцией при P <0,10.
РЕЗУЛЬТАТЫ
Все свиньи остались здоровыми и без проблем потребляли свой рацион. Фекалии были успешно собраны у всех свиней. ATTD AEE был выше при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими рацион CNTR ( P <0.01; ). Однако на ATTD DM, CP, OM, NDF и ADF в рационах не влияло содержание липидов у растущих свиней.
Таблица 3.
ATTD нутриентов в экспериментальных диетах
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
Коэффициенты усвояемости,%
DM
87.3
87,7
88,4
88,2
90,2
89,4
0,78
0,13
OM
89,1
89,4
89,9
90,1
91,5
90,7
0,71 900 0,18
GE
87,7 b
88,4 ab
89,3 ab
89,8 ab
90,9 a
90.4 ab
0,74
0,04
CP
86,2
85,0
85,7
86,4
88,5
86,3
1,19
0,46
AEE 2 900 b
78,5 a
81,5 a
82,7 a
84,5 a
84,2 a
1,59
<0.01
NDF
70,3
67,6
69,2
68,4
73,3
70,3
2,40
0,63
ADF
62,7
61,9
65,3
66,9 9,1 900
61,9
3,72
0,73
Содержание сырого протеина в диете CNTR было выше, чем в липидной диете, что приводит к большему ( P <0.01) потребление азота и удержание азота у свиней, получавших диету CNTR, по сравнению с свиньями, получавшими липидную диету. Кроме того, свиньи в текущем эксперименте получали аналогичное потребление ME (2300 кДж / кг BW 0,6 / день), поэтому свиньи, получавшие рацион CNTR, имели большее ( P <0,01) потребление DM по сравнению со свиньями, получавшими липидные диеты (). Энергия, потерянная в виде мочи для 6 диет, колебалась от 1,5% до 2,1% ДЭ. По сравнению с диетой CNTR, процент потери DE диеты через CH 4 имел тенденцию быть ниже ( P = 0.06) для липидных диет. Отношение NE к ME среди 6 диет колебалось от 77,2% до 80,6%.
Таблица 4.
Влияние характеристик рациона на энергетический и азотный баланс растущих свиней (на основе СВ)
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
BW, кг
41.67
40,14
40,50
42,40
40,95
41,04
1,10
0,74
Потребление DM, кг / сут
1,40 a
1,23 b
1,23 b
1,23 b
1,23 b
1,24 b
1,23 b
1,21 b
0,02
<0,01
Азотный баланс, г / сут
Впуск
38.9 a
30,4 b
28,0 b
29,6 b
29,0 b
28,7 b
0,71
<0,01
0,71
<0,01
a
4,6 ab
4,0 ab
4,0 ab
3,4 b
3,9 ab
0,38
0,02
Выход мочи3 a
5,3 ab
4,8 ab
4,4 b
4,8 ab
5,5 ab
0,61
0,04
a Удержание 26
20,6 b
19,2 b
21,2 b
20,8 b
19,4 b
0,90
<0,01
900 Энергия мочи,% DE
2.1
1,8
1,7
1,5
2,1
1,8
0,20
0,45
Энергия метана,% DE
0,7
0,5
0,6
0,4
0,7
0,5
0,07
0,06
ME / DE
97,2
97,7
97,6
97,7
97,1
97,7
0,29
0,19
NE / ME
77.2 b
80,3 a
80,4 a
80,6 a
80,4 a
80,3 a
0,76
<0,01
кг BW 0,6 / d
ME потребление,
2,284
2,357
2,332
2,341
2,355
2,343
21,95
0,22
Общее производство тепла
9
a
1,276 b
1,287 b
1,261 b
1,266 b
1,280 b
12.52
<0,01
Окисление углеводов 2
1,610 a
1,313 b
1,265 b
1,261 b
1,2476 b
1,2476 b
37,13
<0,01
Окисление жира 3
-417 b
-159 a
-95 a
-111 925
———— а
-90 а
58.02
<0,01
Общая выработка тепла 4 скорректирована
1,380 a
1,245 b
1,269 b
1,238 b
1,236 9000 900 1,256 b
12,50
<0,01
RE P 5
417 a
335 b
311 b
333 2 b
333 2 б
311 б
11.60
<0,01
RE L 6
496 b
746 a
731 a
746 a
751 2
0 a
24,67
<0,01
Сохранение энергии
913 b
1,081 a
1,042 a
1079 a
1085 a
1085
25.24
<0,01
Производство тепла натощак
819
808
790
802
814
819
11,18
0,44
Состояние дыхания
1,10 a
1,04 b
1,03 b
1,03 b
1,02 b
1,03 b
0.01
<0,01
Быстрое состояние
0,82
0,83
0,82
0,83
0,82
0,82
0,01
0,77
Значения энергии, МДж / кг
74 900 GE
18,09
20,52
20,52
20,55
20,63
20,68
—
—
DE
15,86 b
18.14 а
18,25 а
18,32 а
18,57 а
18,59 а
0,14
<0,01
0,14
<0,01
ME
9295 a
17,82 a
17,90 a
18,03 a
18,16 a
0,15
<0,01
NE
11.90 b
14,23 a
14,33 a
14,43 a
14,49 a
14,58 a
0,17
0,17
0,17
свиней, получавших 6 рационов в ходе текущего эксперимента, составляло 2284, 2357, 2332, 2341, 2355 и 2343 кДж / кг BW 0,6 / день, соответственно, что аналогично ожидаемым значениям для плана эксперимента.Добавление диетического жира, независимо от источника, снизило средний THP и скорректированный THP по сравнению со свиньями, получавшими диету CNTR ( P <0,01). Средние скорректированные THP для 6 диет составляли 1380, 1245, 1269, 1238, 1236 и 1256 кДж / кг BW 0,6 / день, соответственно. Тем не менее, средний FHP составлял 809 кДж / кг BW 0,6 · сут -1 для всех обработок и не зависел от характеристик диеты. У свиней, получавших основной корм в сытом состоянии, было больше ( P <0.01) OXCHO для энергии, чем липидные диеты. RE и RE L были выше при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими рацион CNTR ( P <0,01). Кроме того, добавление диетического жира, независимо от источника, уменьшало RQ в сытом состоянии по сравнению со свиньями, получавшими диету CNTR ( P <0,01), но на средний RQ в состоянии натощак не влияли характеристики диеты.
Не было различий по каким-либо измеренным параметрам сывороточного антиоксиданта (SOD, GSH-PX, MDA, T-AOC, CAT и соотношение GSSH / T-GSH) между 6 группами лечения ().По 5 источникам диетического жира, протестированным в нашем исследовании, определенное соотношение DE к GE составляло от 91% до 98%, определенное отношение ME к DE составляло в среднем 99%, определенное отношение NE к ME составляло в среднем 90%, а определенные значения NE колебалась от 32,42 до 34,12 МДж / кг DM. В количественном отношении пальмовое масло имело более низкие значения DE, ME и NE по сравнению с жиром домашней птицы, рыбьим жиром, кукурузным маслом и льняным маслом ().
Таблица 5.
Влияние характеристик рациона на антиоксидантные параметры сыворотки у растущих свиней
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
Параметры антиоксиданта 2
SOD, ед. / Мл
69
130,38
122,80
123,60
126,11
127,47
4,98
0,66
GSH-PX, Ед / мл
681,67
686,17
654,93
611,52 900,9 634,14
20,45
0,11
МДА, нмоль / мл
5,18
4,13
5,16
4,66
4,73
4,31
0.49
0,59
T-AOC, Ед / мл
8,81
8,62
8,29
9,71
8,77
9,09
0,43
0,54
CAT, Ед / мл
5,79
4,87
4,82
4,87
5,31
5,10
0,42
0,55
GSSH / T-GSH
0,23
0,22
0,23
0,22
0.21
0,25
0,01
0,73
Таблица 6.
Использование энергии и содержание энергии в источнике диетического жира у растущих свиней (на основе СВ)
Элемент
Источник липидов
Пальмовое масло
Жир птицы
Рыбий жир
Кукурузное масло
Льняное масло
Использование энергии 1 ,%
DEc / GEc
91.0
93,4
94,4
98,1
97,5
MEc / DEc
99,5
99,2
99,5
96,9
99,3
NEc / MEc
90,6
90,8
91,5
90,1
89,5
Энергетическая ценность 2 , МДж / кг DM
GEm
39,54
39,43
39,33
39,69
39.36
DE
35,98
36,84
37,11
38,95
38,38
ME
35,79
36,56
36,92
37,73
38,11
NE2
38,11
NE2
33,21
33,77
34,00
34,12
ОБСУЖДЕНИЕ
Усвояемость питательных веществ, энергия и баланс азота
Улучшение ATTD AEE для 5 липидных диет в результате увеличения количества пищевых жиров согласуется с предыдущие наблюдения на растущих свиньях (Jørgensen and Fernandez, 2000; Li et al., 2018), предполагая, что эндогенные потери липидов, связанные с потреблением DM корма, оказывают более сильное влияние на ATTD AEE при низких уровнях рациона, чем на более высоких уровнях, а добавленные липиды имеют большую усвояемость, чем липиды из основного рациона (Jørgensen et al., 1993; Noblet, Perez, 1993; Kil et al., 2010).
Удержание азота было ниже при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими CNTR, в основном из-за более низкого суточного потребления азота и аминокислот.Базальная диета, используемая в текущем исследовании, имела гораздо более высокое соотношение белок / ME, чем диеты с добавлением источников липидов, и можно предположить, что только базальная диета используется для поддержания и набора белка и жира. Жир, включенный в экспериментальную диету, в основном способствует отложению, в результате чего в нашем исследовании в виде жира сохранялось на 50% больше энергии. Следовательно, наш экспериментальный план должен обеспечивать значение NE, которое близко к истинному использованию жира в полноценном рационе. Никаких эффектов источника липидов на RE P и RE L не наблюдалось среди 5 липидных диет.Предыдущая работа с крысами и цыплятами-бройлерами показала, что рационы, обогащенные полиненасыщенными жирными кислотами ( PUFA, ), имеют меньшее отложение общего жира в организме, чем рационы, содержащие насыщенные жирные кислоты (Shimomura et al., 1990; Sanz et al., 2000). Их результаты показали, что крысы и цыплята-бройлеры, получавшие рацион, богатый ПНЖК, имели более высокий уровень катаболизма жиров. Кроме того, эндогенный синтез жира (липогенез) может быть снижен у крыс и цыплят-бройлеров, получавших ненасыщенные жирные кислоты. Однако Чвалибог и Торбек (2000) продемонстрировали, что при нормальных условиях кормления основным источником энергии для растущих свиней являются пищевые углеводы, за которыми следует белок.Углеводы и белок обычно могут обеспечить достаточно энергии, чтобы удовлетворить потребности в поддержании и росте, поэтому диетический жир не должен окисляться, но может задерживаться в организме. Это могло быть одной из возможных причин, объясняющих, почему нам не удалось обнаружить другую картину отложения жира у растущих свиней. В сытом состоянии OXF в этом исследовании был отрицательным, что также указывало на то, что свиньи в сытом состоянии не использовали жир для снабжения энергией, а основным субстратом окисления был углевод (Chwalibog et al., 1992).
Добавление диетического жира, независимо от источника, снизило скорректированный THP по сравнению со свиньями, получавшими диету CNTR. Данные, полученные при выращивании свиней в других исследованиях, также подтверждают наш вывод (Noblet et al., 2001; Li et al., 2018). Это наблюдение связано с более высокой эффективностью использования ME из липидов с преимуществом более низкого приращения тепла по сравнению с эффективностью использования ME из белка, крахмала или волокна (Noblet et al., 1994). Другая причина снижения скорректированного THP может заключаться в том, что HP, связанное с активностью кормления, уменьшается, когда липиды добавляются в рационы, скармливаемые растущим свиньям, что приводит к снижению HP (Li et al., 2018). Однако в нашем исследовании физическая активность не измерялась, и поэтому мы не можем проверить эту гипотезу. Даже без количественной оценки физической активности, FHP в нашем испытании измеряли при минимальной ожидаемой активности (ночью, в темноте, регулировке клетки и т. Д.) И после длительного периода голодания. FHP, измеренный в текущем исследовании, составил в среднем 809 кДж / кг BW 0,6 / день, что близко к значениям FHP, полученным в нашей предыдущей работе с использованием того же метода (Liu et al., 2014; Li et al., 2017), а также близкие к тем, которые были получены группой INRA в соответствии с определенной методологией или использовались в крупномасштабных измерениях NE Noblet et al. (1994). Кроме того, в соответствии с другими литературными данными (Le Bellego et al., 2001; Li et al., 2018), состав диеты не влиял на FHP. Лопес Ботэ и др. (2001) предположили, что ненасыщенные жирные кислоты могут увеличивать окислительный стресс у свиней, что может повышать уровень FHP. Однако в этом исследовании параметры диеты не влияли на параметры сывороточного антиоксиданта.Уменьшение RQ для 5 липидных диет в результате увеличения количества пищевых жиров также согласуется с предыдущими наблюдениями на растущих свиньях (Bruininx et al., 2011; Li et al., 2018). С биохимической точки зрения можно ожидать более высокого RQ в CNTR, потому что RQ для синтеза липидов из глюкозы в значительной степени способствует увеличению общего RQ, тогда как отложение диетического жира в виде липидов тела не влияет на общий RQ (van Milgen , 2002).
Энергетическое содержание дополнительных липидов
В NRC (2012) содержание DE в различных источниках липидов основано на исследованиях Wiseman et al.(1990) и Powles et al. (1993, 1994, 1995) с использованием отношения ненасыщенных жирных кислот к насыщенным жирным кислотам и содержания свободных жирных кислот. Значение ME липидов было оценено как 98% от DE, а значение NE было оценено как 88% от ME на основании исследований van Milgen et al. (2001). Оценки ME и NE в NRC (2012) предполагают, что коэффициент преобразования DE в NE одинаков для всех источников жира. Однако было подчеркнуто, что необходимы дополнительные исследования, чтобы определить, верны ли эти отношения и могут ли они широко применяться к различным источникам жира (Kellner and Patience, 2017).По 5 источникам диетического жира, протестированным в нашем исследовании, отношение DE к GE варьировалось от 91% до 98%, в то время как отношение ME к DE (в среднем 99%) и отношение NE к ME (в среднем 90%) были относительно стабильными среди 5 источников липидов. Несмотря на то, что энергетическая эффективность от DE до NE, которую мы определили, больше, чем оценки NRC (2012), наши результаты по отношениям ME / DE и NE / ME полностью согласуются с результатами Noblet et al. (1994), van Milgen et al. (2001) и Ли и др. (2018). Boyd et al. (2015) также пришли к выводу, что энергетический КПД от DE до NE должен быть выше, чем указано в NRC (2012).У других видов, таких как птица, сообщалось о синергетическом эффекте жира с более высоким расчетным значением ME, чем значение GE, когда жир добавлялся в основной рацион (Cao and Adeola, 2016), что можно объяснить лучшим использованием питательных веществ в рационе. базальная диета при наличии жиров. Однако не было найдено литературы, в которой сообщалось бы о перевариваемости жировой энергии выше 100% у растущих свиней.
Точная оценка энергетической ценности пищевых источников жира не только позволяет производителям свинины оценить финансовую ценность дополнительных липидов, но также поддерживает дифференциацию доступных источников жира (Van Heugten et al., 2015; Келлнер и Пейшенс, 2017). Kerr et al. (2015) также подчеркнули необходимость точного определения содержания NE в пищевых липидах, поскольку системы NE могут позволить более точное прогнозирование энергетической ценности пищевых липидов, чем системы DE и ME (Noblet et al., 1994). Несколько исследователей использовали метод сравнительного убоя для определения значения NE липидов (Galloway and Ewan, 1989; Kil et al., 2011), но значения NE, указанные в их исследованиях (17,49, 20,19 и 25,05 МДж / кг сухого вещества, для жир, соевое масло и отборная белая смазка соответственно) были намного меньше, чем оценки Sauvant et al.(2004) и NRC (2012). Однако по 5 источникам диетического жира, протестированным в этом исследовании, средний определенный NE диетического жира составлял 33,50 МДж / кг сухого вещества, а содержание NE в пальмовом масле было численно самым низким (32,42 МДж / кг сухого вещества) в связи с его самым низким показателем. Отношение DE к GE. Это можно объяснить большим количеством насыщенных жирных кислот в пальмовом масле, которые менее растворимы при воздействии солей желчных кислот и могут уменьшать их включение в смешанные мицеллы и замедлять последующее всасывание (Stahly, 1984; Wiseman et al., 1986). Различные составы жирных кислот не имели численных различий в значениях NE для жира птицы, рыбьего жира, кукурузного масла и льняного масла в этом исследовании. Теоретически, когда в рацион добавляют большое количество насыщенных масел, значительная часть пищевых жирных кислот откладывается непосредственно в тканях организма с энергетической эффективностью 90%. Хотя включены большие количества полиненасыщенных масел, некоторые жирные кислоты могут окисляться с выделением энергии в форме АТФ с энергетической эффективностью 66% (Black, 1995; Kil et al., 2011). Однако Ли и др. (2018) предположили, что большая часть пищевых липидов откладывается непосредственно в жировой ткани без каких-либо биохимических превращений после абсорбции, что также подтверждается Bruininx et al. (2011). Эта гипотеза объясняет отсутствие числовой разницы между NE для жира птицы, рыбьего жира, кукурузного масла и льняного масла и относительно высокое отношение NE к ME для всех липидов, которые использовались в этом эксперименте.
Причина основных расхождений между IC и CS при определении содержания NE в липидах в пище заключается в том, что удержание энергии, измеренное с помощью IC, как правило, больше, чем измеренное с помощью CS (Quiniou et al., 1995). В исследованиях с использованием CS свиньи содержатся в более практичных помещениях, а затем могут свободно передвигаться с большим количеством HP, связанных с физической активностью. Кроме того, свиньи в исследованиях с использованием CS могут иногда подниматься ниже своей критической температуры с снова повышенным HP. Следовательно, HP, измеренное с помощью CS, может быть сильно увеличено в связи с увеличением энергии для поддержания и значений FHP (Verstegen, 2001), в то время как значения FHP, измеренные в нашем исследовании с использованием IC, были получены при минимальной активности, при термонейтральности и т. Д.С этой точки зрения результаты с использованием IC должны быть более надежными. Более того, разные стратегии кормления, генетический фон и возраст свиней между нашим исследованием и исследованием Kil et al. (2011) все они могут влиять на содержание, рост и состав тела свиней, что приводит к вариациям в значениях NE липидов (Boisen and Verstegen, 1998). Чтобы подтвердить значение NE пищевых липидов, Boyd et al. (2015) использовали анализ роста для определения NE выбора белой смазки и предположили, что содержание NE в жирах составляет не менее 32.55 МДж / кг для растущих свиней, что убедительно подтверждает наш текущий результат. Очевидно, что необходимы дальнейшие исследования, чтобы определить НЭ липидов в широком диапазоне разнообразных источников пищевых жиров.
ВЫВОДЫ
Значения NE составили 32,42, 33,21, 33,77, 34,00 и 34,12 МДж / кг сухого вещества для пальмового масла, жира птицы, рыбьего жира, кукурузного масла и льняного масла соответственно. Основываясь на наших результатах, отношения ME / DE (приблизительно 99%) и NE / ME (приблизительно 90%) относительно постоянны для всех источников жира, которые мы определили, в то время как соотношение DE / GE более вариабельно среди различных источников жира у растущих свиней. .Для лучшего использования разнообразных источников жира в свиноводстве необходимо больше работать над усвояемостью жира.
Определение содержания чистой энергии в пищевых липидах, скармливаемых растущим свиньям, с помощью непрямой калориметрии
Abstract
Целью этого эксперимента было определение содержания NE в различных пищевых липидах, скармливаемых растущим свиньям, с помощью непрямой калориметрии. Тридцать шесть выращиваемых (исходный BW: 41,1 ± 3,1 кг) курганов были распределены по 6 рационам на основе полностью рандомизированного плана с 6 повторными поросятами на рацион.Диеты включали базальную диету из кукурузо-соевого шрота и 5 тестируемых диет, каждая из которых содержала 10% пальмового масла, птичьего жира, рыбьего жира, кукурузного масла или льняного масла за счет кукурузы и соевого шрота. В течение каждого периода свиней отдельно помещали в клетки для метаболизма на 14 дней, включая 7 дней для адаптации к корму, клеткам для метаболизма и условиям окружающей среды. На 8 день свиней переводили в дыхательные камеры открытого цикла и кормили 1 из 6 рационов из расчета 2,3 МДж ME / кг BW 0,6 / день. Был собран общий кал и моча, а также рассчитана ежедневная выработка тепла ( HP ) с 9 по 13 день.В последний день каждого периода (день 14) свиней голодали и измеряли выработку тепла натощак ( FHP ). Результаты показывают, что FHP свиней в среднем составляла 809 кДж / кг BW 0,6 · день -1 и не зависела от характеристик рациона. Значения DE составляли 35,98, 36,84, 37,11, 38,95 и 38,38 МДж / кг сухого вещества, значения ME составляли 35,79, 36,56, 36,92, 37,73 и 38,11 МДж / кг сухого вещества, а значения NE составляли 32,42, 33,21, 33,77, 34,00 и 34,12 МДж / кг сухого вещества для пальмового масла, жира домашней птицы, рыбьего жира, кукурузного масла и льняного масла соответственно.Основываясь на нашем результате, мы пришли к выводу, что содержание DE диетического липида варьировалось от 91% до 98% от его содержания GE, содержание ME диетического липида составляло приблизительно 99% от его содержания DE, а содержание NE в диетическом липиде было приблизительно 90% его МЭ содержится в растущих свиньях.
Ключевые слова: пищевых липидов, растущие свиньи, косвенная калориметрия, чистая энергия
ВВЕДЕНИЕ
Липиды, концентрированный источник энергии, обычно добавляют в рацион свиней, а энергетическая ценность липидов составляет около 2.В 25 раз больше, чем углеводов (Jones et al., 1992; Lin et al., 2013). Однако энергетическая ценность варьируется в зависимости от источника липидов из-за разного состава жирных кислот, который может влиять на переваривание, абсорбцию и метаболическое использование пищевых липидов (Mendoza and van Heugten, 2014). Также было высказано предположение, что содержание NE в липидах является более точным параметром, чем содержание DE или ME, для описания их продуктивной ценности (Van Heugten et al., 2015). Тем не менее, Sauvant et al. (2004) предложили единое значение NE для всех источников жира (29.77 МДж / кг DM). В NRC (2012) значение NE липидов было оценено в 88% от ME на основании исследования van Milgen et al. (2001). Подчеркивалось, что необходимо более точно определять содержание НЭ в пищевых липидах (Kerr et al., 2015).
Несколько исследователей определили значение NE липидов, используя метод сравнительного убоя ( CS ) (Galloway and Ewan, 1989; Kil et al., 2011). Значение NE для таллового, соевого масла и отборной белой смазки, о которых сообщалось в их исследованиях, было намного меньше, чем у Sauvant et al.(2004) и NRC (2012). Сравнительный метод убоя, который использовал 179 ккал на кг метаболической массы тела (кг 0,6 ) в качестве NE для поддержания (NEm), возможно, занижал значение NE липидов, обогащенных ненасыщенными жирными кислотами, поскольку NEm, вероятно, будет выше в условиях загона и в связь с повышенным окислительным стрессом (López Bote et al., 2001; Verstegen, 2001). Измерение антиоксидантных параметров может помочь эффективно оценить энергетическую ценность пищевых липидов (Shurson et al., 2015).
Таким образом, целью этого эксперимента было определение содержания NE в источниках липидов с различным составом жирных кислот при скармливании растущим свиньям с использованием непрямой калориметрии ( IC ).
МАТЕРИАЛЫ И МЕТОДЫ
Комитет по контролю за благополучием животных и экспериментальной этикой животных лаборатории Китайского сельскохозяйственного университета (Пекин, Китай) рассмотрел и одобрил все протоколы, использованные в этом эксперименте.
Оборудование для непрямой калориметрии
Шесть респираторных камер открытого цикла, использованные в этом эксперименте, были ранее описаны Liu et al.(2014) и Ли и др. (2017). Температуру поддерживали на уровне 22 ° C в сытом состоянии и постепенно повышали до 24 ° C во время голодания. Относительная влажность поддерживалась на уровне 70%. Содержание кислорода измеряли с помощью парамагнитного дифференциального анализатора (Oxymat 6E, Siemens, Мюнхен, Германия), тогда как содержание CO 2 и CH 4 измеряли с помощью инфракрасных анализаторов (Ultramat 6E, Siemens). Анализаторы имели диапазон измерений от 19,5% до 21% для O 2 , от 0% до 1% для CO 2 и от 0% до 0.1% для CH 4 с чувствительностью 0,2% в пределах диапазона измерения. Концентрации газа в каждой камере измерялись с 5-минутными интервалами. Мы использовали эксперимент по сжиганию этанола, чтобы проверить точность камеры при измерении газообмена.
Животные, рационы и план эксперимента
Это исследование проводилось в Исследовательском отделении свиней Феннинг Китайского сельскохозяйственного университета (Хэбэй, Китай). Тридцать шесть растущих курганов (дюрок × ландрас × йоркшир) с начальной BW 41.1 ± 3,1 кг были выделены на 1 из 6 рационов в соответствии с полностью рандомизированным планом с 6 повторениями свиней на рацион. Каждый раз использовали 6 свиней и 6 дыхательных камер открытого цикла, что дало 6 повторных периодов. Рыбий жир и жир домашней птицы, использованные в этом эксперименте, были кормового качества (свободные жирные кислоты <5%), в то время как все остальные липиды были пищевого качества (свободные жирные кислоты <0,2%). Профиль жирных кислот источников липидов представлен в. Диеты включали базальную диету из кукурузно-соевого шрота ( CNTR ) и 5 тестовых диет, каждая из которых содержала 10% пальмового масла ( PALM ), жир птицы ( POUF ), рыбий жир ( FISH ), кукурузное масло (). CORN ) или льняное масло ( FLAX ) за счет кукурузного и соевого шрота ().Все источники липидов были предоставлены компанией Zhongda Agricultural Science and Technology Co., Ltd. (провинция Шаньдун, Китай). Синтезированный лизин, метионин, треонин, триптофан и валин были добавлены, чтобы они превышали потребности животных во все периоды (NRC, 2012).
Таблица 1.
Анализируемый химический состав источников липидов (%, в исходном состоянии)
Элемент
Пальмовое масло
Жир птицы
Рыбий жир
Кукурузное масло
Льняное масло
Жирные кислоты (% от общего количества жирных кислот)
C8: 0
0.1
0,1
0,1
0,1
0,1
C12: 0
0,1
0,1
0,2
0,0
0,0
C14: 0
0,9
0,8
7,0
0,0
0,1
C16: 0
46,2
23,0
19,8
12,7
5,2
C16: 1
0,1
2,7
7.3
0,1
0,1
C17: 0
0,1
0,2
1,5
0,1
0,1
C18: 0
5,1
8,1
4,2
1,9
3,7
C18: 1 n-9
37,2
39,6
15,0
31,4
28,9
C18: 2 n-6
9,1
21,9
2,0
51.9
16,3
C18: 3 n-6
0,0
0,1
0,4
0,0
0,0
C18: 3 n-3
0,4
1,6
1,8
0,7
43,1
C20: 0
0,4
0,3
0,9
0,4
0,3
C20: 1
0,1
0,5
3,1
0,3
0.8
C20: 2
0,0
0,0
0,1
0,0
0,0
C20: 3 n-3
ND 5
0,1
0,2
ND
0,1
C20: 3 n-6
ND
0,1
0,2
ND
0,0
C20: 4 n-6
ND
0,1
1,3
ND
0,0
C20: 5 n-3
ND
0.0
12,4
ND
ND
C21: 0
0,0
0,3
0,3
0,0
0,1
C22: 0
0,1
0,2
0,3
0,0
0,2
C22: 1 n-9
0,0
0,0
0,4
0,0
0,8
C22: 2
ND
0,0
0,1
0.0
0,0
C22: 6 n-3
ND
0,0
18,7
ND
ND
C23: 0
0,0
0,0
0,5
0,0
0,0
C24: 0
0,1
0,1
1,5
0,2
0,1
C24: 1
0,0
0,0
0,7
0,0
0,1
PUFA 1
9.4
23,9
37,1
52,7
59,5
УФА 2
46,9
66,8
63,7
84,5
90,1
SFA 3
53,1
36,2
15,4
9,8
U: S 4
0,9
2,0
1,8
5,5
9,2
Таблица 2.
Ингредиентный состав и химический анализ экспериментальных рационов (%, в исходном состоянии)
Элемент
Диетическое лечение 1
CNTR
PALM
POUF
FISH
CORN
ЛЕН
Состав,%
Кукуруза
70,37
63,13
63,13
63,13
63,13
63.13
Шрот соевый
26,00
23,32
23,32
23,32
23,32
23,32
Пальмовое масло
—
10,00
—
—
—
— 900
Жир птицы
—
—
10,00
—
—
—
Рыбий жир
—
—
—
10.00
—
—
Кукурузное масло
—
—
—
—
10.00
—
Льняное масло
—
—
—
—
10,00
Дикальцийфосфат
1,20
1,20
1,20
1,20
1,20
1,20
Известняк
0.80
0,80
0,80
0,80
0,80
0,80
Соль
0,35
0,35
0,35
0,35
0,35
0,35
Премикс 2 900
0,50
0,50
0,50
0,50
0,50
Lys-HCl 3
0,38
0,34
0,34
0.34
0,34
0,34
DL-Met
0,16
0,14
0,14
0,14
0,14
0,14
L-Thr
0,14
0,13
0,13
0,13
0,13
0,13
L-Trp
0,02
0,02
0,02
0,02
0,02
0,02
L-Val
0.08
0,07
0,07
0,07
0,07
0,07
Анализируемый состав,%
DM
90,07
90,70
90,31
90,36
90,53
89,65
CP
17,49
15,61
15,06
15,01
15,45
15,02
AEE 4
3,83
12.98
12,97
12,52
12,66
12,93
NDF
15,52
13,93
13,67
13,55
13,60
13,83
ADF
3,994 9009 4,4 9009 9,99
3,89
3,66
3,55
Ясень
4,58
4,52
4,44
4,33
4,44
4,39
В течение каждого периода свиньи содержались индивидуально в ящиках для метаболизма в течение 14 d, который включал 7 дней для адаптации к экспериментальной диете, клеткам метаболизма и условиям окружающей среды.На 8 день свиней переводили в дыхательные камеры открытого цикла и кормили 1 из 6 рационов из расчета 2,3 МДж ME / кг BW 0,6 / день. Собирали общий кал и мочу, а также измеряли суточную выработку тепла ( HP ) с 9 по 13 день. В последний день каждого периода (день 14) свиней голодали и производили тепло натощак ( FHP ). измеряли с 22:00 (14-е сутки) до 06:00 (15-е сутки). Период FHP начинался через 30 часов после последнего приема пищи и у животных, которых содержали в темноте, чтобы минимизировать их физическую активность.
Перед экспериментом свиньям скармливали стандартную кукурузно-соевую муку. Свиньи кормили порциями равного размера два раза в день в 09:00 и 16:00. Вода поступала непрерывно через ниппельную поилку низкого давления. Камеры открывали примерно на 1 час в день для кормления свиней и сбора фекалий и мочи. Потребление O 2 и производство CO 2 и CH 4 за это время не были включены в расчет суточного HP. Концентрация CO 2 в камере увеличивалась при закрытии двери.Расчет HP начинался, когда концентрация CO 2 в камере превышала 2000 ppm (Li et al., 2017). Для каждого периода свиней взвешивали на 1, 9, 13 и 14 день. Среднюю массу тела для каждого периода использовали для расчета HP.
Сбор проб
В течение дня с 9 по 13 отбирали остатки корма и просыпи дважды в день для сушки и взвешивания. Общий кал и мочу собирали в соответствии с методами, описанными Liu et al. (2014). Кал собирали дважды в день в 09:00 и 16:00, когда дверь камеры открывали, и сразу хранили при -20 ° C.Мочу собирали отдельно от каждой свиньи в пластиковые ведра, содержащие 50 мл 6 н. HCl, и просеивали через хлопчатобумажную марлю. Был измерен общий объем мочи, произведенной каждой свиньей, и 5% суточной экскреции с мочой сохранялось при -20 ° C. Мочу также собирали отдельно во время 24-часового голодания для расчета потерь азота с мочой для расчета FHP. В конце эксперимента образцы кала и мочи размораживали и тщательно перемешивали, а суб-образец сохраняли для химического анализа.Образцы фекалий сушили в печи в течение 72 часов при 65 ° C. Перед химическим анализом пробы корма и фекалий измельчали через сито диаметром 1 мм. Утром 14-го дня образцы крови (8 мл) были собраны путем пункции передней полой вены с использованием пробирки Vacutainer, не содержащей антикоагулянтов (Becton Dickinson Vacutainer Systems, Franklin Lakes, NJ). Сыворотку получали после центрифугирования (Biofuge22R; Heraeus, Ханау, Германия) при 3000 × g в течение 15 мин при 4 ° C, а затем хранили при -80 ° C до анализа.
Химический анализ
Все химические анализы проводились в двух экземплярах и повторялись, если дубликаты отличались более чем на 5%.Образцы корма и фекалий анализировали на DM (AOAC, 2007; метод 934.01), золу (AOAC 2007; метод 942.15), CP (AOAC, 2007; метод 990.03) и кислотно-гидролизованный эфирный экстракт ( AEE , AOAC, 2007; Метод 954.02). Концентрации NDF и ADF определяли с использованием фильтровальных мешков и оборудования для анализа волокон (Fiber Analyzer, Ankom Technology, Macedon, NY) после модификации процедуры Van Soest et al. (1991). ГЭ в источниках липидов, диетах, образцах мочи и фекалий определяли с помощью калориметра изоперибол (Parr 6300 Calorimeter, Moline, IL) с бензойной кислотой в качестве стандарта.Профили жирных кислот в источниках липидов определяли с помощью газовой хроматографии (серия 6890, Agilent Technologies, Wilmington, DE) по методикам Sukhija и Palmquist (1988) с небольшими изменениями. Образцы липидов были преобразованы в метиловые эфиры жирных кислот с использованием метанольной HCl. Ундекановую кислоту (C11: 0) использовали в качестве внутреннего стандарта. Аликвоты по 1 мкл вводили в капиллярную колонку (60 м × 250 мкм × 250 нм, DB-23, Agilent) с цианопропилметилсиликоном в качестве стационарной фазы.Температуру термостата колонки запрограммировали с разделением 1:20. Температуры инжектора и детектора поддерживались на уровне 260 и 270 ° C соответственно. Азот был газом-носителем при скорости потока 2 мл / мин. Образцы сыворотки размораживали и тщательно перемешивали непосредственно перед анализом. Антиоксидантные параметры, включая супероксиддисмутазу ( SOD ), глутатионпероксидазу ( GSH-PX ), малоновый альдегид ( MDA ), общую антиоксидантную способность ( T-AOC ), каталазу ( CAT ) и соотношение окисленный глутатион ( GSSH ) до общего глутатиона ( T-GSH ) в сыворотке определяли с использованием наборов для анализа в соответствии с инструкциями Beijing Kangjia Bioengineering Company (Пекин, Китай).
Расчеты
Потребление сухого вещества с 9 по 13 день в каждый период рассчитывали как произведение потребления корма и содержания сухого вещества в рационах. Общее потребление энергии рассчитывалось как произведение содержания ГЭ в рационе и фактического потребления сухого вещества корма в течение 5-дневного периода сбора с 9 по 13 день. Энергетические потери с фекалиями, мочой и метаном измерялись для каждого рациона. рассчитывались DE и ME для 6 диет. В ME учитывались потери энергии как с мочой, так и с метаном.Энергия метана была рассчитана с использованием коэффициента преобразования 39,54 кДж / л (Brouwer, 1965).
С 9 по 13 день каждого периода, концентрации O 2 , CO 2 и CH 4 как в входящем, так и в выходящем воздухе, а также скорости выходящего потока воздуха измерялись с 5-минутными интервалами. Эти данные затем использовались для расчета потребления O 2 и производства CO 2 и CH 4 в течение каждого 5-минутного интервала, и эти значения были усреднены и экстраполированы на 24-часовой период.Затем рассчитывалась общая выработка тепла ( THP ) для каждого дня на основе газообмена и потери азота с мочой согласно Брауэру (1965) с использованием уравнения. [1] :):
THP (кДж) = 16,18 × O2 (L) + 5,02 × CO2 (L) — 2,17 × Ch5 (L) — 5,99 × N (г) в моче
(1)
Тепло натощак продуктивность рассчитывалась с использованием того же уравнения, но 24-часовой FHP был спрогнозирован из 8-часового HP после периода отсутствия корма от 31 до 38 часов (с 22:00 до 06:00 часов) в течение последнего дня каждого периода ( Ли и др., 2018). Для базового производства за тот же промежуток времени, что и для расчета THP, 8-часовое HP было также экстраполировано на 24-часовой период. Дыхательный коэффициент ( RQ ) соответствует соотношению между производством CO 2 и потреблением O 2 .
Окисление углеводов ( OXCHO ) и жира ( OXF ) рассчитывали по методу, описанному Chwalibog et al. (1992).
OXCHO (кДж) = [−2,968 × O 2 + 4,147 × CO 2 — 1.761 × CH 4 — 2,446 × N (г) в моче] × 17,58
(2)
OXF (кДж) = [1,719 × O 2 — 1,719 × CO 2 — 1,719 × CH 4 — 1,963 × N (г) в моче] × 39,76
(3)
Кажущуюся общую усвояемость в тракте (ATTD) питательных веществ в рационах рассчитывали в соответствии с методами Noblet et al. (1994).
Сохранение энергии ( RE ) было рассчитано по формуле. [4]:
RE (МДж / кг DM) = [потребление ME (МДж / день) — THP (МДж / день)] / потребление DM (кг / день)
(4)
Сохранение энергии в виде белка ( RE P ) рассчитывали как удерживание N (г) × 6.25 × 23,86 (кДж / г). Сохранение энергии в виде липидов ( RE L ) рассчитывали как разницу между RE и RE P .
Чистая энергия каждого рациона рассчитывалась согласно Noblet et al. (1994) как:
GEc, DEc, Значения MEc и NEc источников липидов были рассчитаны с использованием обновленных методов Adeola (2001) следующим образом:
E липид = [E липидная диета — E базовая диета /96.84 % × 85,70 % ] / 11,19 %
(6)
, где E базальная диета — это среднее значение GE, DE, ME или NE в основной диете (MJ / кг DM ) и соотношение E базальный рацион / 96,84% — это значение GE, DE, ME или NE ингредиентов, которые обеспечивали энергию в базовом рационе. Процент минералов и витаминов в основной диете составлял 3,16% (в пересчете на сухую массу) и не являлся источником энергии.Таким образом, ингредиенты, обеспечивающие энергию (кукуруза, SBM и АК), составляют 96,84% в базовом рационе на основе сухого вещества. Предполагалось, что значения GE, DE, ME и NE смеси кукурузы, SBM и AA, полученные из основного рациона, применимы к другим диетам. E липидная диета — среднее значение GE, DE, ME или NE в липидной диете (МДж / кг СВ). E липид — это расчетные значения GE, DE, ME и NE ( GEc , DEc , MEc, и NEc ) для каждого образца липидов (МДж / кг DM).85,70% — это процент DM ингредиентов (кроме липидов), которые поставляли энергию (кукуруза, SBM и AA) в липидной диете, и 11,19% — это процент DM в образце липидов в липидной диете. DM ингредиентов измеряли перед приготовлением диет, чтобы рассчитать соотношение DM каждого тестируемого ингредиента в рационе. Отношения DEc / GEc, MEc / Dec и NEc / MEc затем могут быть рассчитаны для каждого источника липидов на основе этих рассчитанных значений GEc, DEc, MEc и NEc и использованы для оценки конечных значений DE, ME и NE в качестве продукта. измеренных GEm и DEc / GEc для DE, измеренных GEm и DEc / GEc и MEc / DEc для ME и измеренных GEm и DEc / GEc, MEc / DEc и NEc / MEc для NE (Li et al., 2017). Все расчеты производились на основе DM. Поскольку существует только одно оценочное значение энергии для каждого источника липидов, статистический анализ не может быть проведен среди значений энергии источников липидов.
Статистический анализ
Данные были проверены на нормальность, и выбросы были обнаружены с использованием процедуры UNIVARIATE SAS (Институт SAS, Кэри, Северная Каролина). Никаких выбросов не выявлено. Затем данные были проанализированы с использованием ANOVA с процедурой MIXED SAS. Отдельную свинью рассматривали как экспериментальную единицу, и статистическая модель включала лечебную диету как единственный фиксированный эффект, а период и камеру как случайные эффекты.Средние значения лечения были рассчитаны с использованием инструкции LSMEANS, а статистические различия между видами лечения были разделены с использованием теста Tukey HSD. Во всех анализах различия считались значимыми, если P <0,05, и считались тенденцией при P <0,10.
РЕЗУЛЬТАТЫ
Все свиньи остались здоровыми и без проблем потребляли свой рацион. Фекалии были успешно собраны у всех свиней. ATTD AEE был выше при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими рацион CNTR ( P <0.01; ). Однако на ATTD DM, CP, OM, NDF и ADF в рационах не влияло содержание липидов у растущих свиней.
Таблица 3.
ATTD нутриентов в экспериментальных диетах
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
Коэффициенты усвояемости,%
DM
87.3
87,7
88,4
88,2
90,2
89,4
0,78
0,13
OM
89,1
89,4
89,9
90,1
91,5
90,7
0,71 900 0,18
GE
87,7 b
88,4 ab
89,3 ab
89,8 ab
90,9 a
90.4 ab
0,74
0,04
CP
86,2
85,0
85,7
86,4
88,5
86,3
1,19
0,46
AEE 2 900 b
78,5 a
81,5 a
82,7 a
84,5 a
84,2 a
1,59
<0.01
NDF
70,3
67,6
69,2
68,4
73,3
70,3
2,40
0,63
ADF
62,7
61,9
65,3
66,9 9,1 900
61,9
3,72
0,73
Содержание сырого протеина в диете CNTR было выше, чем в липидной диете, что приводит к большему ( P <0.01) потребление азота и удержание азота у свиней, получавших диету CNTR, по сравнению с свиньями, получавшими липидную диету. Кроме того, свиньи в текущем эксперименте получали аналогичное потребление ME (2300 кДж / кг BW 0,6 / день), поэтому свиньи, получавшие рацион CNTR, имели большее ( P <0,01) потребление DM по сравнению со свиньями, получавшими липидные диеты (). Энергия, потерянная в виде мочи для 6 диет, колебалась от 1,5% до 2,1% ДЭ. По сравнению с диетой CNTR, процент потери DE диеты через CH 4 имел тенденцию быть ниже ( P = 0.06) для липидных диет. Отношение NE к ME среди 6 диет колебалось от 77,2% до 80,6%.
Таблица 4.
Влияние характеристик рациона на энергетический и азотный баланс растущих свиней (на основе СВ)
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
BW, кг
41.67
40,14
40,50
42,40
40,95
41,04
1,10
0,74
Потребление DM, кг / сут
1,40 a
1,23 b
1,23 b
1,23 b
1,23 b
1,24 b
1,23 b
1,21 b
0,02
<0,01
Азотный баланс, г / сут
Впуск
38.9 a
30,4 b
28,0 b
29,6 b
29,0 b
28,7 b
0,71
<0,01
0,71
<0,01
a
4,6 ab
4,0 ab
4,0 ab
3,4 b
3,9 ab
0,38
0,02
Выход мочи3 a
5,3 ab
4,8 ab
4,4 b
4,8 ab
5,5 ab
0,61
0,04
a Удержание 26
20,6 b
19,2 b
21,2 b
20,8 b
19,4 b
0,90
<0,01
900 Энергия мочи,% DE
2.1
1,8
1,7
1,5
2,1
1,8
0,20
0,45
Энергия метана,% DE
0,7
0,5
0,6
0,4
0,7
0,5
0,07
0,06
ME / DE
97,2
97,7
97,6
97,7
97,1
97,7
0,29
0,19
NE / ME
77.2 b
80,3 a
80,4 a
80,6 a
80,4 a
80,3 a
0,76
<0,01
кг BW 0,6 / d
ME потребление,
2,284
2,357
2,332
2,341
2,355
2,343
21,95
0,22
Общее производство тепла
9
a
1,276 b
1,287 b
1,261 b
1,266 b
1,280 b
12.52
<0,01
Окисление углеводов 2
1,610 a
1,313 b
1,265 b
1,261 b
1,2476 b
1,2476 b
37,13
<0,01
Окисление жира 3
-417 b
-159 a
-95 a
-111 925
———— а
-90 а
58.02
<0,01
Общая выработка тепла 4 скорректирована
1,380 a
1,245 b
1,269 b
1,238 b
1,236 9000 900 1,256 b
12,50
<0,01
RE P 5
417 a
335 b
311 b
333 2 b
333 2 б
311 б
11.60
<0,01
RE L 6
496 b
746 a
731 a
746 a
751 2
0 a
24,67
<0,01
Сохранение энергии
913 b
1,081 a
1,042 a
1079 a
1085 a
1085
25.24
<0,01
Производство тепла натощак
819
808
790
802
814
819
11,18
0,44
Состояние дыхания
1,10 a
1,04 b
1,03 b
1,03 b
1,02 b
1,03 b
0.01
<0,01
Быстрое состояние
0,82
0,83
0,82
0,83
0,82
0,82
0,01
0,77
Значения энергии, МДж / кг
74 900 GE
18,09
20,52
20,52
20,55
20,63
20,68
—
—
DE
15,86 b
18.14 а
18,25 а
18,32 а
18,57 а
18,59 а
0,14
<0,01
0,14
<0,01
ME
9295 a
17,82 a
17,90 a
18,03 a
18,16 a
0,15
<0,01
NE
11.90 b
14,23 a
14,33 a
14,43 a
14,49 a
14,58 a
0,17
0,17
0,17
свиней, получавших 6 рационов в ходе текущего эксперимента, составляло 2284, 2357, 2332, 2341, 2355 и 2343 кДж / кг BW 0,6 / день, соответственно, что аналогично ожидаемым значениям для плана эксперимента.Добавление диетического жира, независимо от источника, снизило средний THP и скорректированный THP по сравнению со свиньями, получавшими диету CNTR ( P <0,01). Средние скорректированные THP для 6 диет составляли 1380, 1245, 1269, 1238, 1236 и 1256 кДж / кг BW 0,6 / день, соответственно. Тем не менее, средний FHP составлял 809 кДж / кг BW 0,6 · сут -1 для всех обработок и не зависел от характеристик диеты. У свиней, получавших основной корм в сытом состоянии, было больше ( P <0.01) OXCHO для энергии, чем липидные диеты. RE и RE L были выше при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими рацион CNTR ( P <0,01). Кроме того, добавление диетического жира, независимо от источника, уменьшало RQ в сытом состоянии по сравнению со свиньями, получавшими диету CNTR ( P <0,01), но на средний RQ в состоянии натощак не влияли характеристики диеты.
Не было различий по каким-либо измеренным параметрам сывороточного антиоксиданта (SOD, GSH-PX, MDA, T-AOC, CAT и соотношение GSSH / T-GSH) между 6 группами лечения ().По 5 источникам диетического жира, протестированным в нашем исследовании, определенное соотношение DE к GE составляло от 91% до 98%, определенное отношение ME к DE составляло в среднем 99%, определенное отношение NE к ME составляло в среднем 90%, а определенные значения NE колебалась от 32,42 до 34,12 МДж / кг DM. В количественном отношении пальмовое масло имело более низкие значения DE, ME и NE по сравнению с жиром домашней птицы, рыбьим жиром, кукурузным маслом и льняным маслом ().
Таблица 5.
Влияние характеристик рациона на антиоксидантные параметры сыворотки у растущих свиней
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
Параметры антиоксиданта 2
SOD, ед. / Мл
69
130,38
122,80
123,60
126,11
127,47
4,98
0,66
GSH-PX, Ед / мл
681,67
686,17
654,93
611,52 900,9 634,14
20,45
0,11
МДА, нмоль / мл
5,18
4,13
5,16
4,66
4,73
4,31
0.49
0,59
T-AOC, Ед / мл
8,81
8,62
8,29
9,71
8,77
9,09
0,43
0,54
CAT, Ед / мл
5,79
4,87
4,82
4,87
5,31
5,10
0,42
0,55
GSSH / T-GSH
0,23
0,22
0,23
0,22
0.21
0,25
0,01
0,73
Таблица 6.
Использование энергии и содержание энергии в источнике диетического жира у растущих свиней (на основе СВ)
Элемент
Источник липидов
Пальмовое масло
Жир птицы
Рыбий жир
Кукурузное масло
Льняное масло
Использование энергии 1 ,%
DEc / GEc
91.0
93,4
94,4
98,1
97,5
MEc / DEc
99,5
99,2
99,5
96,9
99,3
NEc / MEc
90,6
90,8
91,5
90,1
89,5
Энергетическая ценность 2 , МДж / кг DM
GEm
39,54
39,43
39,33
39,69
39.36
DE
35,98
36,84
37,11
38,95
38,38
ME
35,79
36,56
36,92
37,73
38,11
NE2
38,11
NE2
33,21
33,77
34,00
34,12
ОБСУЖДЕНИЕ
Усвояемость питательных веществ, энергия и баланс азота
Улучшение ATTD AEE для 5 липидных диет в результате увеличения количества пищевых жиров согласуется с предыдущие наблюдения на растущих свиньях (Jørgensen and Fernandez, 2000; Li et al., 2018), предполагая, что эндогенные потери липидов, связанные с потреблением DM корма, оказывают более сильное влияние на ATTD AEE при низких уровнях рациона, чем на более высоких уровнях, а добавленные липиды имеют большую усвояемость, чем липиды из основного рациона (Jørgensen et al., 1993; Noblet, Perez, 1993; Kil et al., 2010).
Удержание азота было ниже при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими CNTR, в основном из-за более низкого суточного потребления азота и аминокислот.Базальная диета, используемая в текущем исследовании, имела гораздо более высокое соотношение белок / ME, чем диеты с добавлением источников липидов, и можно предположить, что только базальная диета используется для поддержания и набора белка и жира. Жир, включенный в экспериментальную диету, в основном способствует отложению, в результате чего в нашем исследовании в виде жира сохранялось на 50% больше энергии. Следовательно, наш экспериментальный план должен обеспечивать значение NE, которое близко к истинному использованию жира в полноценном рационе. Никаких эффектов источника липидов на RE P и RE L не наблюдалось среди 5 липидных диет.Предыдущая работа с крысами и цыплятами-бройлерами показала, что рационы, обогащенные полиненасыщенными жирными кислотами ( PUFA, ), имеют меньшее отложение общего жира в организме, чем рационы, содержащие насыщенные жирные кислоты (Shimomura et al., 1990; Sanz et al., 2000). Их результаты показали, что крысы и цыплята-бройлеры, получавшие рацион, богатый ПНЖК, имели более высокий уровень катаболизма жиров. Кроме того, эндогенный синтез жира (липогенез) может быть снижен у крыс и цыплят-бройлеров, получавших ненасыщенные жирные кислоты. Однако Чвалибог и Торбек (2000) продемонстрировали, что при нормальных условиях кормления основным источником энергии для растущих свиней являются пищевые углеводы, за которыми следует белок.Углеводы и белок обычно могут обеспечить достаточно энергии, чтобы удовлетворить потребности в поддержании и росте, поэтому диетический жир не должен окисляться, но может задерживаться в организме. Это могло быть одной из возможных причин, объясняющих, почему нам не удалось обнаружить другую картину отложения жира у растущих свиней. В сытом состоянии OXF в этом исследовании был отрицательным, что также указывало на то, что свиньи в сытом состоянии не использовали жир для снабжения энергией, а основным субстратом окисления был углевод (Chwalibog et al., 1992).
Добавление диетического жира, независимо от источника, снизило скорректированный THP по сравнению со свиньями, получавшими диету CNTR. Данные, полученные при выращивании свиней в других исследованиях, также подтверждают наш вывод (Noblet et al., 2001; Li et al., 2018). Это наблюдение связано с более высокой эффективностью использования ME из липидов с преимуществом более низкого приращения тепла по сравнению с эффективностью использования ME из белка, крахмала или волокна (Noblet et al., 1994). Другая причина снижения скорректированного THP может заключаться в том, что HP, связанное с активностью кормления, уменьшается, когда липиды добавляются в рационы, скармливаемые растущим свиньям, что приводит к снижению HP (Li et al., 2018). Однако в нашем исследовании физическая активность не измерялась, и поэтому мы не можем проверить эту гипотезу. Даже без количественной оценки физической активности, FHP в нашем испытании измеряли при минимальной ожидаемой активности (ночью, в темноте, регулировке клетки и т. Д.) И после длительного периода голодания. FHP, измеренный в текущем исследовании, составил в среднем 809 кДж / кг BW 0,6 / день, что близко к значениям FHP, полученным в нашей предыдущей работе с использованием того же метода (Liu et al., 2014; Li et al., 2017), а также близкие к тем, которые были получены группой INRA в соответствии с определенной методологией или использовались в крупномасштабных измерениях NE Noblet et al. (1994). Кроме того, в соответствии с другими литературными данными (Le Bellego et al., 2001; Li et al., 2018), состав диеты не влиял на FHP. Лопес Ботэ и др. (2001) предположили, что ненасыщенные жирные кислоты могут увеличивать окислительный стресс у свиней, что может повышать уровень FHP. Однако в этом исследовании параметры диеты не влияли на параметры сывороточного антиоксиданта.Уменьшение RQ для 5 липидных диет в результате увеличения количества пищевых жиров также согласуется с предыдущими наблюдениями на растущих свиньях (Bruininx et al., 2011; Li et al., 2018). С биохимической точки зрения можно ожидать более высокого RQ в CNTR, потому что RQ для синтеза липидов из глюкозы в значительной степени способствует увеличению общего RQ, тогда как отложение диетического жира в виде липидов тела не влияет на общий RQ (van Milgen , 2002).
Энергетическое содержание дополнительных липидов
В NRC (2012) содержание DE в различных источниках липидов основано на исследованиях Wiseman et al.(1990) и Powles et al. (1993, 1994, 1995) с использованием отношения ненасыщенных жирных кислот к насыщенным жирным кислотам и содержания свободных жирных кислот. Значение ME липидов было оценено как 98% от DE, а значение NE было оценено как 88% от ME на основании исследований van Milgen et al. (2001). Оценки ME и NE в NRC (2012) предполагают, что коэффициент преобразования DE в NE одинаков для всех источников жира. Однако было подчеркнуто, что необходимы дополнительные исследования, чтобы определить, верны ли эти отношения и могут ли они широко применяться к различным источникам жира (Kellner and Patience, 2017).По 5 источникам диетического жира, протестированным в нашем исследовании, отношение DE к GE варьировалось от 91% до 98%, в то время как отношение ME к DE (в среднем 99%) и отношение NE к ME (в среднем 90%) были относительно стабильными среди 5 источников липидов. Несмотря на то, что энергетическая эффективность от DE до NE, которую мы определили, больше, чем оценки NRC (2012), наши результаты по отношениям ME / DE и NE / ME полностью согласуются с результатами Noblet et al. (1994), van Milgen et al. (2001) и Ли и др. (2018). Boyd et al. (2015) также пришли к выводу, что энергетический КПД от DE до NE должен быть выше, чем указано в NRC (2012).У других видов, таких как птица, сообщалось о синергетическом эффекте жира с более высоким расчетным значением ME, чем значение GE, когда жир добавлялся в основной рацион (Cao and Adeola, 2016), что можно объяснить лучшим использованием питательных веществ в рационе. базальная диета при наличии жиров. Однако не было найдено литературы, в которой сообщалось бы о перевариваемости жировой энергии выше 100% у растущих свиней.
Точная оценка энергетической ценности пищевых источников жира не только позволяет производителям свинины оценить финансовую ценность дополнительных липидов, но также поддерживает дифференциацию доступных источников жира (Van Heugten et al., 2015; Келлнер и Пейшенс, 2017). Kerr et al. (2015) также подчеркнули необходимость точного определения содержания NE в пищевых липидах, поскольку системы NE могут позволить более точное прогнозирование энергетической ценности пищевых липидов, чем системы DE и ME (Noblet et al., 1994). Несколько исследователей использовали метод сравнительного убоя для определения значения NE липидов (Galloway and Ewan, 1989; Kil et al., 2011), но значения NE, указанные в их исследованиях (17,49, 20,19 и 25,05 МДж / кг сухого вещества, для жир, соевое масло и отборная белая смазка соответственно) были намного меньше, чем оценки Sauvant et al.(2004) и NRC (2012). Однако по 5 источникам диетического жира, протестированным в этом исследовании, средний определенный NE диетического жира составлял 33,50 МДж / кг сухого вещества, а содержание NE в пальмовом масле было численно самым низким (32,42 МДж / кг сухого вещества) в связи с его самым низким показателем. Отношение DE к GE. Это можно объяснить большим количеством насыщенных жирных кислот в пальмовом масле, которые менее растворимы при воздействии солей желчных кислот и могут уменьшать их включение в смешанные мицеллы и замедлять последующее всасывание (Stahly, 1984; Wiseman et al., 1986). Различные составы жирных кислот не имели численных различий в значениях NE для жира птицы, рыбьего жира, кукурузного масла и льняного масла в этом исследовании. Теоретически, когда в рацион добавляют большое количество насыщенных масел, значительная часть пищевых жирных кислот откладывается непосредственно в тканях организма с энергетической эффективностью 90%. Хотя включены большие количества полиненасыщенных масел, некоторые жирные кислоты могут окисляться с выделением энергии в форме АТФ с энергетической эффективностью 66% (Black, 1995; Kil et al., 2011). Однако Ли и др. (2018) предположили, что большая часть пищевых липидов откладывается непосредственно в жировой ткани без каких-либо биохимических превращений после абсорбции, что также подтверждается Bruininx et al. (2011). Эта гипотеза объясняет отсутствие числовой разницы между NE для жира птицы, рыбьего жира, кукурузного масла и льняного масла и относительно высокое отношение NE к ME для всех липидов, которые использовались в этом эксперименте.
Причина основных расхождений между IC и CS при определении содержания NE в липидах в пище заключается в том, что удержание энергии, измеренное с помощью IC, как правило, больше, чем измеренное с помощью CS (Quiniou et al., 1995). В исследованиях с использованием CS свиньи содержатся в более практичных помещениях, а затем могут свободно передвигаться с большим количеством HP, связанных с физической активностью. Кроме того, свиньи в исследованиях с использованием CS могут иногда подниматься ниже своей критической температуры с снова повышенным HP. Следовательно, HP, измеренное с помощью CS, может быть сильно увеличено в связи с увеличением энергии для поддержания и значений FHP (Verstegen, 2001), в то время как значения FHP, измеренные в нашем исследовании с использованием IC, были получены при минимальной активности, при термонейтральности и т. Д.С этой точки зрения результаты с использованием IC должны быть более надежными. Более того, разные стратегии кормления, генетический фон и возраст свиней между нашим исследованием и исследованием Kil et al. (2011) все они могут влиять на содержание, рост и состав тела свиней, что приводит к вариациям в значениях NE липидов (Boisen and Verstegen, 1998). Чтобы подтвердить значение NE пищевых липидов, Boyd et al. (2015) использовали анализ роста для определения NE выбора белой смазки и предположили, что содержание NE в жирах составляет не менее 32.55 МДж / кг для растущих свиней, что убедительно подтверждает наш текущий результат. Очевидно, что необходимы дальнейшие исследования, чтобы определить НЭ липидов в широком диапазоне разнообразных источников пищевых жиров.
ВЫВОДЫ
Значения NE составили 32,42, 33,21, 33,77, 34,00 и 34,12 МДж / кг сухого вещества для пальмового масла, жира птицы, рыбьего жира, кукурузного масла и льняного масла соответственно. Основываясь на наших результатах, отношения ME / DE (приблизительно 99%) и NE / ME (приблизительно 90%) относительно постоянны для всех источников жира, которые мы определили, в то время как соотношение DE / GE более вариабельно среди различных источников жира у растущих свиней. .Для лучшего использования разнообразных источников жира в свиноводстве необходимо больше работать над усвояемостью жира.
Определение содержания чистой энергии в пищевых липидах, скармливаемых растущим свиньям, с помощью непрямой калориметрии
Abstract
Целью этого эксперимента было определение содержания NE в различных пищевых липидах, скармливаемых растущим свиньям, с помощью непрямой калориметрии. Тридцать шесть выращиваемых (исходный BW: 41,1 ± 3,1 кг) курганов были распределены по 6 рационам на основе полностью рандомизированного плана с 6 повторными поросятами на рацион.Диеты включали базальную диету из кукурузо-соевого шрота и 5 тестируемых диет, каждая из которых содержала 10% пальмового масла, птичьего жира, рыбьего жира, кукурузного масла или льняного масла за счет кукурузы и соевого шрота. В течение каждого периода свиней отдельно помещали в клетки для метаболизма на 14 дней, включая 7 дней для адаптации к корму, клеткам для метаболизма и условиям окружающей среды. На 8 день свиней переводили в дыхательные камеры открытого цикла и кормили 1 из 6 рационов из расчета 2,3 МДж ME / кг BW 0,6 / день. Был собран общий кал и моча, а также рассчитана ежедневная выработка тепла ( HP ) с 9 по 13 день.В последний день каждого периода (день 14) свиней голодали и измеряли выработку тепла натощак ( FHP ). Результаты показывают, что FHP свиней в среднем составляла 809 кДж / кг BW 0,6 · день -1 и не зависела от характеристик рациона. Значения DE составляли 35,98, 36,84, 37,11, 38,95 и 38,38 МДж / кг сухого вещества, значения ME составляли 35,79, 36,56, 36,92, 37,73 и 38,11 МДж / кг сухого вещества, а значения NE составляли 32,42, 33,21, 33,77, 34,00 и 34,12 МДж / кг сухого вещества для пальмового масла, жира домашней птицы, рыбьего жира, кукурузного масла и льняного масла соответственно.Основываясь на нашем результате, мы пришли к выводу, что содержание DE диетического липида варьировалось от 91% до 98% от его содержания GE, содержание ME диетического липида составляло приблизительно 99% от его содержания DE, а содержание NE в диетическом липиде было приблизительно 90% его МЭ содержится в растущих свиньях.
Ключевые слова: пищевых липидов, растущие свиньи, косвенная калориметрия, чистая энергия
ВВЕДЕНИЕ
Липиды, концентрированный источник энергии, обычно добавляют в рацион свиней, а энергетическая ценность липидов составляет около 2.В 25 раз больше, чем углеводов (Jones et al., 1992; Lin et al., 2013). Однако энергетическая ценность варьируется в зависимости от источника липидов из-за разного состава жирных кислот, который может влиять на переваривание, абсорбцию и метаболическое использование пищевых липидов (Mendoza and van Heugten, 2014). Также было высказано предположение, что содержание NE в липидах является более точным параметром, чем содержание DE или ME, для описания их продуктивной ценности (Van Heugten et al., 2015). Тем не менее, Sauvant et al. (2004) предложили единое значение NE для всех источников жира (29.77 МДж / кг DM). В NRC (2012) значение NE липидов было оценено в 88% от ME на основании исследования van Milgen et al. (2001). Подчеркивалось, что необходимо более точно определять содержание НЭ в пищевых липидах (Kerr et al., 2015).
Несколько исследователей определили значение NE липидов, используя метод сравнительного убоя ( CS ) (Galloway and Ewan, 1989; Kil et al., 2011). Значение NE для таллового, соевого масла и отборной белой смазки, о которых сообщалось в их исследованиях, было намного меньше, чем у Sauvant et al.(2004) и NRC (2012). Сравнительный метод убоя, который использовал 179 ккал на кг метаболической массы тела (кг 0,6 ) в качестве NE для поддержания (NEm), возможно, занижал значение NE липидов, обогащенных ненасыщенными жирными кислотами, поскольку NEm, вероятно, будет выше в условиях загона и в связь с повышенным окислительным стрессом (López Bote et al., 2001; Verstegen, 2001). Измерение антиоксидантных параметров может помочь эффективно оценить энергетическую ценность пищевых липидов (Shurson et al., 2015).
Таким образом, целью этого эксперимента было определение содержания NE в источниках липидов с различным составом жирных кислот при скармливании растущим свиньям с использованием непрямой калориметрии ( IC ).
МАТЕРИАЛЫ И МЕТОДЫ
Комитет по контролю за благополучием животных и экспериментальной этикой животных лаборатории Китайского сельскохозяйственного университета (Пекин, Китай) рассмотрел и одобрил все протоколы, использованные в этом эксперименте.
Оборудование для непрямой калориметрии
Шесть респираторных камер открытого цикла, использованные в этом эксперименте, были ранее описаны Liu et al.(2014) и Ли и др. (2017). Температуру поддерживали на уровне 22 ° C в сытом состоянии и постепенно повышали до 24 ° C во время голодания. Относительная влажность поддерживалась на уровне 70%. Содержание кислорода измеряли с помощью парамагнитного дифференциального анализатора (Oxymat 6E, Siemens, Мюнхен, Германия), тогда как содержание CO 2 и CH 4 измеряли с помощью инфракрасных анализаторов (Ultramat 6E, Siemens). Анализаторы имели диапазон измерений от 19,5% до 21% для O 2 , от 0% до 1% для CO 2 и от 0% до 0.1% для CH 4 с чувствительностью 0,2% в пределах диапазона измерения. Концентрации газа в каждой камере измерялись с 5-минутными интервалами. Мы использовали эксперимент по сжиганию этанола, чтобы проверить точность камеры при измерении газообмена.
Животные, рационы и план эксперимента
Это исследование проводилось в Исследовательском отделении свиней Феннинг Китайского сельскохозяйственного университета (Хэбэй, Китай). Тридцать шесть растущих курганов (дюрок × ландрас × йоркшир) с начальной BW 41.1 ± 3,1 кг были выделены на 1 из 6 рационов в соответствии с полностью рандомизированным планом с 6 повторениями свиней на рацион. Каждый раз использовали 6 свиней и 6 дыхательных камер открытого цикла, что дало 6 повторных периодов. Рыбий жир и жир домашней птицы, использованные в этом эксперименте, были кормового качества (свободные жирные кислоты <5%), в то время как все остальные липиды были пищевого качества (свободные жирные кислоты <0,2%). Профиль жирных кислот источников липидов представлен в. Диеты включали базальную диету из кукурузно-соевого шрота ( CNTR ) и 5 тестовых диет, каждая из которых содержала 10% пальмового масла ( PALM ), жир птицы ( POUF ), рыбий жир ( FISH ), кукурузное масло (). CORN ) или льняное масло ( FLAX ) за счет кукурузного и соевого шрота ().Все источники липидов были предоставлены компанией Zhongda Agricultural Science and Technology Co., Ltd. (провинция Шаньдун, Китай). Синтезированный лизин, метионин, треонин, триптофан и валин были добавлены, чтобы они превышали потребности животных во все периоды (NRC, 2012).
Таблица 1.
Анализируемый химический состав источников липидов (%, в исходном состоянии)
Элемент
Пальмовое масло
Жир птицы
Рыбий жир
Кукурузное масло
Льняное масло
Жирные кислоты (% от общего количества жирных кислот)
C8: 0
0.1
0,1
0,1
0,1
0,1
C12: 0
0,1
0,1
0,2
0,0
0,0
C14: 0
0,9
0,8
7,0
0,0
0,1
C16: 0
46,2
23,0
19,8
12,7
5,2
C16: 1
0,1
2,7
7.3
0,1
0,1
C17: 0
0,1
0,2
1,5
0,1
0,1
C18: 0
5,1
8,1
4,2
1,9
3,7
C18: 1 n-9
37,2
39,6
15,0
31,4
28,9
C18: 2 n-6
9,1
21,9
2,0
51.9
16,3
C18: 3 n-6
0,0
0,1
0,4
0,0
0,0
C18: 3 n-3
0,4
1,6
1,8
0,7
43,1
C20: 0
0,4
0,3
0,9
0,4
0,3
C20: 1
0,1
0,5
3,1
0,3
0.8
C20: 2
0,0
0,0
0,1
0,0
0,0
C20: 3 n-3
ND 5
0,1
0,2
ND
0,1
C20: 3 n-6
ND
0,1
0,2
ND
0,0
C20: 4 n-6
ND
0,1
1,3
ND
0,0
C20: 5 n-3
ND
0.0
12,4
ND
ND
C21: 0
0,0
0,3
0,3
0,0
0,1
C22: 0
0,1
0,2
0,3
0,0
0,2
C22: 1 n-9
0,0
0,0
0,4
0,0
0,8
C22: 2
ND
0,0
0,1
0.0
0,0
C22: 6 n-3
ND
0,0
18,7
ND
ND
C23: 0
0,0
0,0
0,5
0,0
0,0
C24: 0
0,1
0,1
1,5
0,2
0,1
C24: 1
0,0
0,0
0,7
0,0
0,1
PUFA 1
9.4
23,9
37,1
52,7
59,5
УФА 2
46,9
66,8
63,7
84,5
90,1
SFA 3
53,1
36,2
15,4
9,8
U: S 4
0,9
2,0
1,8
5,5
9,2
Таблица 2.
Ингредиентный состав и химический анализ экспериментальных рационов (%, в исходном состоянии)
Элемент
Диетическое лечение 1
CNTR
PALM
POUF
FISH
CORN
ЛЕН
Состав,%
Кукуруза
70,37
63,13
63,13
63,13
63,13
63.13
Шрот соевый
26,00
23,32
23,32
23,32
23,32
23,32
Пальмовое масло
—
10,00
—
—
—
— 900
Жир птицы
—
—
10,00
—
—
—
Рыбий жир
—
—
—
10.00
—
—
Кукурузное масло
—
—
—
—
10.00
—
Льняное масло
—
—
—
—
10,00
Дикальцийфосфат
1,20
1,20
1,20
1,20
1,20
1,20
Известняк
0.80
0,80
0,80
0,80
0,80
0,80
Соль
0,35
0,35
0,35
0,35
0,35
0,35
Премикс 2 900
0,50
0,50
0,50
0,50
0,50
Lys-HCl 3
0,38
0,34
0,34
0.34
0,34
0,34
DL-Met
0,16
0,14
0,14
0,14
0,14
0,14
L-Thr
0,14
0,13
0,13
0,13
0,13
0,13
L-Trp
0,02
0,02
0,02
0,02
0,02
0,02
L-Val
0.08
0,07
0,07
0,07
0,07
0,07
Анализируемый состав,%
DM
90,07
90,70
90,31
90,36
90,53
89,65
CP
17,49
15,61
15,06
15,01
15,45
15,02
AEE 4
3,83
12.98
12,97
12,52
12,66
12,93
NDF
15,52
13,93
13,67
13,55
13,60
13,83
ADF
3,994 9009 4,4 9009 9,99
3,89
3,66
3,55
Ясень
4,58
4,52
4,44
4,33
4,44
4,39
В течение каждого периода свиньи содержались индивидуально в ящиках для метаболизма в течение 14 d, который включал 7 дней для адаптации к экспериментальной диете, клеткам метаболизма и условиям окружающей среды.На 8 день свиней переводили в дыхательные камеры открытого цикла и кормили 1 из 6 рационов из расчета 2,3 МДж ME / кг BW 0,6 / день. Собирали общий кал и мочу, а также измеряли суточную выработку тепла ( HP ) с 9 по 13 день. В последний день каждого периода (день 14) свиней голодали и производили тепло натощак ( FHP ). измеряли с 22:00 (14-е сутки) до 06:00 (15-е сутки). Период FHP начинался через 30 часов после последнего приема пищи и у животных, которых содержали в темноте, чтобы минимизировать их физическую активность.
Перед экспериментом свиньям скармливали стандартную кукурузно-соевую муку. Свиньи кормили порциями равного размера два раза в день в 09:00 и 16:00. Вода поступала непрерывно через ниппельную поилку низкого давления. Камеры открывали примерно на 1 час в день для кормления свиней и сбора фекалий и мочи. Потребление O 2 и производство CO 2 и CH 4 за это время не были включены в расчет суточного HP. Концентрация CO 2 в камере увеличивалась при закрытии двери.Расчет HP начинался, когда концентрация CO 2 в камере превышала 2000 ppm (Li et al., 2017). Для каждого периода свиней взвешивали на 1, 9, 13 и 14 день. Среднюю массу тела для каждого периода использовали для расчета HP.
Сбор проб
В течение дня с 9 по 13 отбирали остатки корма и просыпи дважды в день для сушки и взвешивания. Общий кал и мочу собирали в соответствии с методами, описанными Liu et al. (2014). Кал собирали дважды в день в 09:00 и 16:00, когда дверь камеры открывали, и сразу хранили при -20 ° C.Мочу собирали отдельно от каждой свиньи в пластиковые ведра, содержащие 50 мл 6 н. HCl, и просеивали через хлопчатобумажную марлю. Был измерен общий объем мочи, произведенной каждой свиньей, и 5% суточной экскреции с мочой сохранялось при -20 ° C. Мочу также собирали отдельно во время 24-часового голодания для расчета потерь азота с мочой для расчета FHP. В конце эксперимента образцы кала и мочи размораживали и тщательно перемешивали, а суб-образец сохраняли для химического анализа.Образцы фекалий сушили в печи в течение 72 часов при 65 ° C. Перед химическим анализом пробы корма и фекалий измельчали через сито диаметром 1 мм. Утром 14-го дня образцы крови (8 мл) были собраны путем пункции передней полой вены с использованием пробирки Vacutainer, не содержащей антикоагулянтов (Becton Dickinson Vacutainer Systems, Franklin Lakes, NJ). Сыворотку получали после центрифугирования (Biofuge22R; Heraeus, Ханау, Германия) при 3000 × g в течение 15 мин при 4 ° C, а затем хранили при -80 ° C до анализа.
Химический анализ
Все химические анализы проводились в двух экземплярах и повторялись, если дубликаты отличались более чем на 5%.Образцы корма и фекалий анализировали на DM (AOAC, 2007; метод 934.01), золу (AOAC 2007; метод 942.15), CP (AOAC, 2007; метод 990.03) и кислотно-гидролизованный эфирный экстракт ( AEE , AOAC, 2007; Метод 954.02). Концентрации NDF и ADF определяли с использованием фильтровальных мешков и оборудования для анализа волокон (Fiber Analyzer, Ankom Technology, Macedon, NY) после модификации процедуры Van Soest et al. (1991). ГЭ в источниках липидов, диетах, образцах мочи и фекалий определяли с помощью калориметра изоперибол (Parr 6300 Calorimeter, Moline, IL) с бензойной кислотой в качестве стандарта.Профили жирных кислот в источниках липидов определяли с помощью газовой хроматографии (серия 6890, Agilent Technologies, Wilmington, DE) по методикам Sukhija и Palmquist (1988) с небольшими изменениями. Образцы липидов были преобразованы в метиловые эфиры жирных кислот с использованием метанольной HCl. Ундекановую кислоту (C11: 0) использовали в качестве внутреннего стандарта. Аликвоты по 1 мкл вводили в капиллярную колонку (60 м × 250 мкм × 250 нм, DB-23, Agilent) с цианопропилметилсиликоном в качестве стационарной фазы.Температуру термостата колонки запрограммировали с разделением 1:20. Температуры инжектора и детектора поддерживались на уровне 260 и 270 ° C соответственно. Азот был газом-носителем при скорости потока 2 мл / мин. Образцы сыворотки размораживали и тщательно перемешивали непосредственно перед анализом. Антиоксидантные параметры, включая супероксиддисмутазу ( SOD ), глутатионпероксидазу ( GSH-PX ), малоновый альдегид ( MDA ), общую антиоксидантную способность ( T-AOC ), каталазу ( CAT ) и соотношение окисленный глутатион ( GSSH ) до общего глутатиона ( T-GSH ) в сыворотке определяли с использованием наборов для анализа в соответствии с инструкциями Beijing Kangjia Bioengineering Company (Пекин, Китай).
Расчеты
Потребление сухого вещества с 9 по 13 день в каждый период рассчитывали как произведение потребления корма и содержания сухого вещества в рационах. Общее потребление энергии рассчитывалось как произведение содержания ГЭ в рационе и фактического потребления сухого вещества корма в течение 5-дневного периода сбора с 9 по 13 день. Энергетические потери с фекалиями, мочой и метаном измерялись для каждого рациона. рассчитывались DE и ME для 6 диет. В ME учитывались потери энергии как с мочой, так и с метаном.Энергия метана была рассчитана с использованием коэффициента преобразования 39,54 кДж / л (Brouwer, 1965).
С 9 по 13 день каждого периода, концентрации O 2 , CO 2 и CH 4 как в входящем, так и в выходящем воздухе, а также скорости выходящего потока воздуха измерялись с 5-минутными интервалами. Эти данные затем использовались для расчета потребления O 2 и производства CO 2 и CH 4 в течение каждого 5-минутного интервала, и эти значения были усреднены и экстраполированы на 24-часовой период.Затем рассчитывалась общая выработка тепла ( THP ) для каждого дня на основе газообмена и потери азота с мочой согласно Брауэру (1965) с использованием уравнения. [1] :):
THP (кДж) = 16,18 × O2 (L) + 5,02 × CO2 (L) — 2,17 × Ch5 (L) — 5,99 × N (г) в моче
(1)
Тепло натощак продуктивность рассчитывалась с использованием того же уравнения, но 24-часовой FHP был спрогнозирован из 8-часового HP после периода отсутствия корма от 31 до 38 часов (с 22:00 до 06:00 часов) в течение последнего дня каждого периода ( Ли и др., 2018). Для базового производства за тот же промежуток времени, что и для расчета THP, 8-часовое HP было также экстраполировано на 24-часовой период. Дыхательный коэффициент ( RQ ) соответствует соотношению между производством CO 2 и потреблением O 2 .
Окисление углеводов ( OXCHO ) и жира ( OXF ) рассчитывали по методу, описанному Chwalibog et al. (1992).
OXCHO (кДж) = [−2,968 × O 2 + 4,147 × CO 2 — 1.761 × CH 4 — 2,446 × N (г) в моче] × 17,58
(2)
OXF (кДж) = [1,719 × O 2 — 1,719 × CO 2 — 1,719 × CH 4 — 1,963 × N (г) в моче] × 39,76
(3)
Кажущуюся общую усвояемость в тракте (ATTD) питательных веществ в рационах рассчитывали в соответствии с методами Noblet et al. (1994).
Сохранение энергии ( RE ) было рассчитано по формуле. [4]:
RE (МДж / кг DM) = [потребление ME (МДж / день) — THP (МДж / день)] / потребление DM (кг / день)
(4)
Сохранение энергии в виде белка ( RE P ) рассчитывали как удерживание N (г) × 6.25 × 23,86 (кДж / г). Сохранение энергии в виде липидов ( RE L ) рассчитывали как разницу между RE и RE P .
Чистая энергия каждого рациона рассчитывалась согласно Noblet et al. (1994) как:
GEc, DEc, Значения MEc и NEc источников липидов были рассчитаны с использованием обновленных методов Adeola (2001) следующим образом:
E липид = [E липидная диета — E базовая диета /96.84 % × 85,70 % ] / 11,19 %
(6)
, где E базальная диета — это среднее значение GE, DE, ME или NE в основной диете (MJ / кг DM ) и соотношение E базальный рацион / 96,84% — это значение GE, DE, ME или NE ингредиентов, которые обеспечивали энергию в базовом рационе. Процент минералов и витаминов в основной диете составлял 3,16% (в пересчете на сухую массу) и не являлся источником энергии.Таким образом, ингредиенты, обеспечивающие энергию (кукуруза, SBM и АК), составляют 96,84% в базовом рационе на основе сухого вещества. Предполагалось, что значения GE, DE, ME и NE смеси кукурузы, SBM и AA, полученные из основного рациона, применимы к другим диетам. E липидная диета — среднее значение GE, DE, ME или NE в липидной диете (МДж / кг СВ). E липид — это расчетные значения GE, DE, ME и NE ( GEc , DEc , MEc, и NEc ) для каждого образца липидов (МДж / кг DM).85,70% — это процент DM ингредиентов (кроме липидов), которые поставляли энергию (кукуруза, SBM и AA) в липидной диете, и 11,19% — это процент DM в образце липидов в липидной диете. DM ингредиентов измеряли перед приготовлением диет, чтобы рассчитать соотношение DM каждого тестируемого ингредиента в рационе. Отношения DEc / GEc, MEc / Dec и NEc / MEc затем могут быть рассчитаны для каждого источника липидов на основе этих рассчитанных значений GEc, DEc, MEc и NEc и использованы для оценки конечных значений DE, ME и NE в качестве продукта. измеренных GEm и DEc / GEc для DE, измеренных GEm и DEc / GEc и MEc / DEc для ME и измеренных GEm и DEc / GEc, MEc / DEc и NEc / MEc для NE (Li et al., 2017). Все расчеты производились на основе DM. Поскольку существует только одно оценочное значение энергии для каждого источника липидов, статистический анализ не может быть проведен среди значений энергии источников липидов.
Статистический анализ
Данные были проверены на нормальность, и выбросы были обнаружены с использованием процедуры UNIVARIATE SAS (Институт SAS, Кэри, Северная Каролина). Никаких выбросов не выявлено. Затем данные были проанализированы с использованием ANOVA с процедурой MIXED SAS. Отдельную свинью рассматривали как экспериментальную единицу, и статистическая модель включала лечебную диету как единственный фиксированный эффект, а период и камеру как случайные эффекты.Средние значения лечения были рассчитаны с использованием инструкции LSMEANS, а статистические различия между видами лечения были разделены с использованием теста Tukey HSD. Во всех анализах различия считались значимыми, если P <0,05, и считались тенденцией при P <0,10.
РЕЗУЛЬТАТЫ
Все свиньи остались здоровыми и без проблем потребляли свой рацион. Фекалии были успешно собраны у всех свиней. ATTD AEE был выше при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими рацион CNTR ( P <0.01; ). Однако на ATTD DM, CP, OM, NDF и ADF в рационах не влияло содержание липидов у растущих свиней.
Таблица 3.
ATTD нутриентов в экспериментальных диетах
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
Коэффициенты усвояемости,%
DM
87.3
87,7
88,4
88,2
90,2
89,4
0,78
0,13
OM
89,1
89,4
89,9
90,1
91,5
90,7
0,71 900 0,18
GE
87,7 b
88,4 ab
89,3 ab
89,8 ab
90,9 a
90.4 ab
0,74
0,04
CP
86,2
85,0
85,7
86,4
88,5
86,3
1,19
0,46
AEE 2 900 b
78,5 a
81,5 a
82,7 a
84,5 a
84,2 a
1,59
<0.01
NDF
70,3
67,6
69,2
68,4
73,3
70,3
2,40
0,63
ADF
62,7
61,9
65,3
66,9 9,1 900
61,9
3,72
0,73
Содержание сырого протеина в диете CNTR было выше, чем в липидной диете, что приводит к большему ( P <0.01) потребление азота и удержание азота у свиней, получавших диету CNTR, по сравнению с свиньями, получавшими липидную диету. Кроме того, свиньи в текущем эксперименте получали аналогичное потребление ME (2300 кДж / кг BW 0,6 / день), поэтому свиньи, получавшие рацион CNTR, имели большее ( P <0,01) потребление DM по сравнению со свиньями, получавшими липидные диеты (). Энергия, потерянная в виде мочи для 6 диет, колебалась от 1,5% до 2,1% ДЭ. По сравнению с диетой CNTR, процент потери DE диеты через CH 4 имел тенденцию быть ниже ( P = 0.06) для липидных диет. Отношение NE к ME среди 6 диет колебалось от 77,2% до 80,6%.
Таблица 4.
Влияние характеристик рациона на энергетический и азотный баланс растущих свиней (на основе СВ)
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
BW, кг
41.67
40,14
40,50
42,40
40,95
41,04
1,10
0,74
Потребление DM, кг / сут
1,40 a
1,23 b
1,23 b
1,23 b
1,23 b
1,24 b
1,23 b
1,21 b
0,02
<0,01
Азотный баланс, г / сут
Впуск
38.9 a
30,4 b
28,0 b
29,6 b
29,0 b
28,7 b
0,71
<0,01
0,71
<0,01
a
4,6 ab
4,0 ab
4,0 ab
3,4 b
3,9 ab
0,38
0,02
Выход мочи3 a
5,3 ab
4,8 ab
4,4 b
4,8 ab
5,5 ab
0,61
0,04
a Удержание 26
20,6 b
19,2 b
21,2 b
20,8 b
19,4 b
0,90
<0,01
900 Энергия мочи,% DE
2.1
1,8
1,7
1,5
2,1
1,8
0,20
0,45
Энергия метана,% DE
0,7
0,5
0,6
0,4
0,7
0,5
0,07
0,06
ME / DE
97,2
97,7
97,6
97,7
97,1
97,7
0,29
0,19
NE / ME
77.2 b
80,3 a
80,4 a
80,6 a
80,4 a
80,3 a
0,76
<0,01
кг BW 0,6 / d
ME потребление,
2,284
2,357
2,332
2,341
2,355
2,343
21,95
0,22
Общее производство тепла
9
a
1,276 b
1,287 b
1,261 b
1,266 b
1,280 b
12.52
<0,01
Окисление углеводов 2
1,610 a
1,313 b
1,265 b
1,261 b
1,2476 b
1,2476 b
37,13
<0,01
Окисление жира 3
-417 b
-159 a
-95 a
-111 925
———— а
-90 а
58.02
<0,01
Общая выработка тепла 4 скорректирована
1,380 a
1,245 b
1,269 b
1,238 b
1,236 9000 900 1,256 b
12,50
<0,01
RE P 5
417 a
335 b
311 b
333 2 b
333 2 б
311 б
11.60
<0,01
RE L 6
496 b
746 a
731 a
746 a
751 2
0 a
24,67
<0,01
Сохранение энергии
913 b
1,081 a
1,042 a
1079 a
1085 a
1085
25.24
<0,01
Производство тепла натощак
819
808
790
802
814
819
11,18
0,44
Состояние дыхания
1,10 a
1,04 b
1,03 b
1,03 b
1,02 b
1,03 b
0.01
<0,01
Быстрое состояние
0,82
0,83
0,82
0,83
0,82
0,82
0,01
0,77
Значения энергии, МДж / кг
74 900 GE
18,09
20,52
20,52
20,55
20,63
20,68
—
—
DE
15,86 b
18.14 а
18,25 а
18,32 а
18,57 а
18,59 а
0,14
<0,01
0,14
<0,01
ME
9295 a
17,82 a
17,90 a
18,03 a
18,16 a
0,15
<0,01
NE
11.90 b
14,23 a
14,33 a
14,43 a
14,49 a
14,58 a
0,17
0,17
0,17
свиней, получавших 6 рационов в ходе текущего эксперимента, составляло 2284, 2357, 2332, 2341, 2355 и 2343 кДж / кг BW 0,6 / день, соответственно, что аналогично ожидаемым значениям для плана эксперимента.Добавление диетического жира, независимо от источника, снизило средний THP и скорректированный THP по сравнению со свиньями, получавшими диету CNTR ( P <0,01). Средние скорректированные THP для 6 диет составляли 1380, 1245, 1269, 1238, 1236 и 1256 кДж / кг BW 0,6 / день, соответственно. Тем не менее, средний FHP составлял 809 кДж / кг BW 0,6 · сут -1 для всех обработок и не зависел от характеристик диеты. У свиней, получавших основной корм в сытом состоянии, было больше ( P <0.01) OXCHO для энергии, чем липидные диеты. RE и RE L были выше при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими рацион CNTR ( P <0,01). Кроме того, добавление диетического жира, независимо от источника, уменьшало RQ в сытом состоянии по сравнению со свиньями, получавшими диету CNTR ( P <0,01), но на средний RQ в состоянии натощак не влияли характеристики диеты.
Не было различий по каким-либо измеренным параметрам сывороточного антиоксиданта (SOD, GSH-PX, MDA, T-AOC, CAT и соотношение GSSH / T-GSH) между 6 группами лечения ().По 5 источникам диетического жира, протестированным в нашем исследовании, определенное соотношение DE к GE составляло от 91% до 98%, определенное отношение ME к DE составляло в среднем 99%, определенное отношение NE к ME составляло в среднем 90%, а определенные значения NE колебалась от 32,42 до 34,12 МДж / кг DM. В количественном отношении пальмовое масло имело более низкие значения DE, ME и NE по сравнению с жиром домашней птицы, рыбьим жиром, кукурузным маслом и льняным маслом ().
Таблица 5.
Влияние характеристик рациона на антиоксидантные параметры сыворотки у растущих свиней
Элемент
Диетическое лечение 1
SEM
P -значение
CNTR
PALM
POUF
FISH
CORN
FLAX
Параметры антиоксиданта 2
SOD, ед. / Мл
69
130,38
122,80
123,60
126,11
127,47
4,98
0,66
GSH-PX, Ед / мл
681,67
686,17
654,93
611,52 900,9 634,14
20,45
0,11
МДА, нмоль / мл
5,18
4,13
5,16
4,66
4,73
4,31
0.49
0,59
T-AOC, Ед / мл
8,81
8,62
8,29
9,71
8,77
9,09
0,43
0,54
CAT, Ед / мл
5,79
4,87
4,82
4,87
5,31
5,10
0,42
0,55
GSSH / T-GSH
0,23
0,22
0,23
0,22
0.21
0,25
0,01
0,73
Таблица 6.
Использование энергии и содержание энергии в источнике диетического жира у растущих свиней (на основе СВ)
Элемент
Источник липидов
Пальмовое масло
Жир птицы
Рыбий жир
Кукурузное масло
Льняное масло
Использование энергии 1 ,%
DEc / GEc
91.0
93,4
94,4
98,1
97,5
MEc / DEc
99,5
99,2
99,5
96,9
99,3
NEc / MEc
90,6
90,8
91,5
90,1
89,5
Энергетическая ценность 2 , МДж / кг DM
GEm
39,54
39,43
39,33
39,69
39.36
DE
35,98
36,84
37,11
38,95
38,38
ME
35,79
36,56
36,92
37,73
38,11
NE2
38,11
NE2
33,21
33,77
34,00
34,12
ОБСУЖДЕНИЕ
Усвояемость питательных веществ, энергия и баланс азота
Улучшение ATTD AEE для 5 липидных диет в результате увеличения количества пищевых жиров согласуется с предыдущие наблюдения на растущих свиньях (Jørgensen and Fernandez, 2000; Li et al., 2018), предполагая, что эндогенные потери липидов, связанные с потреблением DM корма, оказывают более сильное влияние на ATTD AEE при низких уровнях рациона, чем на более высоких уровнях, а добавленные липиды имеют большую усвояемость, чем липиды из основного рациона (Jørgensen et al., 1993; Noblet, Perez, 1993; Kil et al., 2010).
Удержание азота было ниже при добавлении диетического жира, независимо от источника, по сравнению со свиньями, получавшими CNTR, в основном из-за более низкого суточного потребления азота и аминокислот.Базальная диета, используемая в текущем исследовании, имела гораздо более высокое соотношение белок / ME, чем диеты с добавлением источников липидов, и можно предположить, что только базальная диета используется для поддержания и набора белка и жира. Жир, включенный в экспериментальную диету, в основном способствует отложению, в результате чего в нашем исследовании в виде жира сохранялось на 50% больше энергии. Следовательно, наш экспериментальный план должен обеспечивать значение NE, которое близко к истинному использованию жира в полноценном рационе. Никаких эффектов источника липидов на RE P и RE L не наблюдалось среди 5 липидных диет.Предыдущая работа с крысами и цыплятами-бройлерами показала, что рационы, обогащенные полиненасыщенными жирными кислотами ( PUFA, ), имеют меньшее отложение общего жира в организме, чем рационы, содержащие насыщенные жирные кислоты (Shimomura et al., 1990; Sanz et al., 2000). Их результаты показали, что крысы и цыплята-бройлеры, получавшие рацион, богатый ПНЖК, имели более высокий уровень катаболизма жиров. Кроме того, эндогенный синтез жира (липогенез) может быть снижен у крыс и цыплят-бройлеров, получавших ненасыщенные жирные кислоты. Однако Чвалибог и Торбек (2000) продемонстрировали, что при нормальных условиях кормления основным источником энергии для растущих свиней являются пищевые углеводы, за которыми следует белок.Углеводы и белок обычно могут обеспечить достаточно энергии, чтобы удовлетворить потребности в поддержании и росте, поэтому диетический жир не должен окисляться, но может задерживаться в организме. Это могло быть одной из возможных причин, объясняющих, почему нам не удалось обнаружить другую картину отложения жира у растущих свиней. В сытом состоянии OXF в этом исследовании был отрицательным, что также указывало на то, что свиньи в сытом состоянии не использовали жир для снабжения энергией, а основным субстратом окисления был углевод (Chwalibog et al., 1992).
Добавление диетического жира, независимо от источника, снизило скорректированный THP по сравнению со свиньями, получавшими диету CNTR. Данные, полученные при выращивании свиней в других исследованиях, также подтверждают наш вывод (Noblet et al., 2001; Li et al., 2018). Это наблюдение связано с более высокой эффективностью использования ME из липидов с преимуществом более низкого приращения тепла по сравнению с эффективностью использования ME из белка, крахмала или волокна (Noblet et al., 1994). Другая причина снижения скорректированного THP может заключаться в том, что HP, связанное с активностью кормления, уменьшается, когда липиды добавляются в рационы, скармливаемые растущим свиньям, что приводит к снижению HP (Li et al., 2018). Однако в нашем исследовании физическая активность не измерялась, и поэтому мы не можем проверить эту гипотезу. Даже без количественной оценки физической активности, FHP в нашем испытании измеряли при минимальной ожидаемой активности (ночью, в темноте, регулировке клетки и т. Д.) И после длительного периода голодания. FHP, измеренный в текущем исследовании, составил в среднем 809 кДж / кг BW 0,6 / день, что близко к значениям FHP, полученным в нашей предыдущей работе с использованием того же метода (Liu et al., 2014; Li et al., 2017), а также близкие к тем, которые были получены группой INRA в соответствии с определенной методологией или использовались в крупномасштабных измерениях NE Noblet et al. (1994). Кроме того, в соответствии с другими литературными данными (Le Bellego et al., 2001; Li et al., 2018), состав диеты не влиял на FHP. Лопес Ботэ и др. (2001) предположили, что ненасыщенные жирные кислоты могут увеличивать окислительный стресс у свиней, что может повышать уровень FHP. Однако в этом исследовании параметры диеты не влияли на параметры сывороточного антиоксиданта.Уменьшение RQ для 5 липидных диет в результате увеличения количества пищевых жиров также согласуется с предыдущими наблюдениями на растущих свиньях (Bruininx et al., 2011; Li et al., 2018). С биохимической точки зрения можно ожидать более высокого RQ в CNTR, потому что RQ для синтеза липидов из глюкозы в значительной степени способствует увеличению общего RQ, тогда как отложение диетического жира в виде липидов тела не влияет на общий RQ (van Milgen , 2002).
Энергетическое содержание дополнительных липидов
В NRC (2012) содержание DE в различных источниках липидов основано на исследованиях Wiseman et al.(1990) и Powles et al. (1993, 1994, 1995) с использованием отношения ненасыщенных жирных кислот к насыщенным жирным кислотам и содержания свободных жирных кислот. Значение ME липидов было оценено как 98% от DE, а значение NE было оценено как 88% от ME на основании исследований van Milgen et al. (2001). Оценки ME и NE в NRC (2012) предполагают, что коэффициент преобразования DE в NE одинаков для всех источников жира. Однако было подчеркнуто, что необходимы дополнительные исследования, чтобы определить, верны ли эти отношения и могут ли они широко применяться к различным источникам жира (Kellner and Patience, 2017).По 5 источникам диетического жира, протестированным в нашем исследовании, отношение DE к GE варьировалось от 91% до 98%, в то время как отношение ME к DE (в среднем 99%) и отношение NE к ME (в среднем 90%) были относительно стабильными среди 5 источников липидов. Несмотря на то, что энергетическая эффективность от DE до NE, которую мы определили, больше, чем оценки NRC (2012), наши результаты по отношениям ME / DE и NE / ME полностью согласуются с результатами Noblet et al. (1994), van Milgen et al. (2001) и Ли и др. (2018). Boyd et al. (2015) также пришли к выводу, что энергетический КПД от DE до NE должен быть выше, чем указано в NRC (2012).У других видов, таких как птица, сообщалось о синергетическом эффекте жира с более высоким расчетным значением ME, чем значение GE, когда жир добавлялся в основной рацион (Cao and Adeola, 2016), что можно объяснить лучшим использованием питательных веществ в рационе. базальная диета при наличии жиров. Однако не было найдено литературы, в которой сообщалось бы о перевариваемости жировой энергии выше 100% у растущих свиней.
Точная оценка энергетической ценности пищевых источников жира не только позволяет производителям свинины оценить финансовую ценность дополнительных липидов, но также поддерживает дифференциацию доступных источников жира (Van Heugten et al., 2015; Келлнер и Пейшенс, 2017). Kerr et al. (2015) также подчеркнули необходимость точного определения содержания NE в пищевых липидах, поскольку системы NE могут позволить более точное прогнозирование энергетической ценности пищевых липидов, чем системы DE и ME (Noblet et al., 1994). Несколько исследователей использовали метод сравнительного убоя для определения значения NE липидов (Galloway and Ewan, 1989; Kil et al., 2011), но значения NE, указанные в их исследованиях (17,49, 20,19 и 25,05 МДж / кг сухого вещества, для жир, соевое масло и отборная белая смазка соответственно) были намного меньше, чем оценки Sauvant et al.(2004) и NRC (2012). Однако по 5 источникам диетического жира, протестированным в этом исследовании, средний определенный NE диетического жира составлял 33,50 МДж / кг сухого вещества, а содержание NE в пальмовом масле было численно самым низким (32,42 МДж / кг сухого вещества) в связи с его самым низким показателем. Отношение DE к GE. Это можно объяснить большим количеством насыщенных жирных кислот в пальмовом масле, которые менее растворимы при воздействии солей желчных кислот и могут уменьшать их включение в смешанные мицеллы и замедлять последующее всасывание (Stahly, 1984; Wiseman et al., 1986). Различные составы жирных кислот не имели численных различий в значениях NE для жира птицы, рыбьего жира, кукурузного масла и льняного масла в этом исследовании. Теоретически, когда в рацион добавляют большое количество насыщенных масел, значительная часть пищевых жирных кислот откладывается непосредственно в тканях организма с энергетической эффективностью 90%. Хотя включены большие количества полиненасыщенных масел, некоторые жирные кислоты могут окисляться с выделением энергии в форме АТФ с энергетической эффективностью 66% (Black, 1995; Kil et al., 2011). Однако Ли и др. (2018) предположили, что большая часть пищевых липидов откладывается непосредственно в жировой ткани без каких-либо биохимических превращений после абсорбции, что также подтверждается Bruininx et al. (2011). Эта гипотеза объясняет отсутствие числовой разницы между NE для жира птицы, рыбьего жира, кукурузного масла и льняного масла и относительно высокое отношение NE к ME для всех липидов, которые использовались в этом эксперименте.
Причина основных расхождений между IC и CS при определении содержания NE в липидах в пище заключается в том, что удержание энергии, измеренное с помощью IC, как правило, больше, чем измеренное с помощью CS (Quiniou et al., 1995). В исследованиях с использованием CS свиньи содержатся в более практичных помещениях, а затем могут свободно передвигаться с большим количеством HP, связанных с физической активностью. Кроме того, свиньи в исследованиях с использованием CS могут иногда подниматься ниже своей критической температуры с снова повышенным HP. Следовательно, HP, измеренное с помощью CS, может быть сильно увеличено в связи с увеличением энергии для поддержания и значений FHP (Verstegen, 2001), в то время как значения FHP, измеренные в нашем исследовании с использованием IC, были получены при минимальной активности, при термонейтральности и т. Д.С этой точки зрения результаты с использованием IC должны быть более надежными. Более того, разные стратегии кормления, генетический фон и возраст свиней между нашим исследованием и исследованием Kil et al. (2011) все они могут влиять на содержание, рост и состав тела свиней, что приводит к вариациям в значениях NE липидов (Boisen and Verstegen, 1998). Чтобы подтвердить значение NE пищевых липидов, Boyd et al. (2015) использовали анализ роста для определения NE выбора белой смазки и предположили, что содержание NE в жирах составляет не менее 32.55 МДж / кг для растущих свиней, что убедительно подтверждает наш текущий результат. Очевидно, что необходимы дальнейшие исследования, чтобы определить НЭ липидов в широком диапазоне разнообразных источников пищевых жиров.
ВЫВОДЫ
Значения NE составили 32,42, 33,21, 33,77, 34,00 и 34,12 МДж / кг сухого вещества для пальмового масла, жира птицы, рыбьего жира, кукурузного масла и льняного масла соответственно. Основываясь на наших результатах, отношения ME / DE (приблизительно 99%) и NE / ME (приблизительно 90%) относительно постоянны для всех источников жира, которые мы определили, в то время как соотношение DE / GE более вариабельно среди различных источников жира у растущих свиней. .Для лучшего использования разнообразных источников жира в свиноводстве необходимо больше работать над усвояемостью жира.
Характеристика липидов и их кормовая ценность в рационах свиней | Журнал зоотехники и биотехнологии
1.
Statisticia. 2014. Statisticia: Статистический портал. http://www.statista.com/ дата обращения 1 мая 2015 г.
2.
NRA. 2014. Национальная ассоциация визуализаторов. http://www.nationalrenderers.org/ дата обращения 1 мая 2015 г.
Палмквист DL. Омега-3 жирные кислоты в обмене веществ, здоровье и питании, а также для модифицированных продуктов животного происхождения. Prof Anim Sci. 2009; 25: 207–49.
Google ученый
14.
Палмквист Д.Л., Дженкинс ТС. Проблемы с методами жиров и жирных кислот. J Anim Sci. 2003. 81: 3250–4.
CAS
PubMed
Google ученый
15.
Лутрия, Д. Л. 2004. Добыча и анализ нефти: основные вопросы и сравнительные исследования.Редактор Д. Л. Лутрии. AOCS Press, Шампейн, Иллинойс.
16.
Моллер Дж. Зерновые, зерновые продукты и корма для животных — определение сырого и общего жира методом экстракции Рэндалла: совместное исследование. Обеспечение качества и безопасности сельскохозяйственных культур и продуктов питания. 2010. с. 1–6.
Google ученый
17.
Jongbloed R, Smits B. Влияние HCl-гидролиза для определения сырого жира на содержание сырого жира, усвояемость сырого жира и NE f кормов для свиней на откорме.Отчет IVVO-DLR № 263.DLO-Институт зоотехники и здоровья (ID-DLO) — филиал Runderweg. 1994.
Google ученый
18.
Шурсон Г.К., Керр Б.Дж., Хансон А.Р. Оценка качества кормовых жиров и масел и их влияния на показатели роста свиней. J Anim Sci Biotech. 2015; 6: 1–11.
CAS
Google ученый
19.
Jones PJH, Rideout T. Липиды, стерины и их метаболиты.В: Росс А.С., Кабальеро Б., Казинс Р.Дж., Такер К.Л., Циглер Т.Р., редакторы. Современное питание в здоровье и болезни. 11 об. Изд. Филадельфия, Пенсильвания: Липпинкотт Уильямс и Уилкинс; 2012. с. 65–87.
Google ученый
20.
Хамош М. Лингвальные и желудочные липазы. Питание. 1990; 6: 421–8.
Ланглуа А., Корринг Т., Левенез Ф., Кубер Дж. К., Чайвиалле Дж. А. Влияние полипептида поджелудочной железы на отток желчных путей и секрецию желчных кислот, стимулируемую секретином и холецистокинином у свиньи в сознании. Regul Pept. 1990; 27: 139–47.
CAS
PubMed
Google ученый
23.
Borgstrom B, Erlanson C.Взаимодействие липазы и колипазы поджелудочной железы и влияние солей желчных кислот и других детергентов. Eur J Biochem. 1973; 37: 60–8.
CAS
PubMed
Google ученый
24.
Vandermeers A, Vandermeers-Piret MC, Rathe J, Christophe J. О триацилглицеринлипазе поджелудочной железы человека: выделение и некоторые свойства. Biochim Biophys Acta. 1974. 370: 257–68.
CAS
PubMed
Google ученый
25.
Borgstrom B. Важность фосфолипидов, панкреатической фосфолипазы A2 и жирных кислот для переваривания пищевых жиров: эксперименты in vivo со свиными ферментами. Гастроэнтерология. 1980; 78: 954–62.
Hoffmann AF, Mekhijian HF. Желчные кислоты и кишечное всасывание жиров и электролитов при здоровье и болезни. В кн .: Наир П.П., Кричевский Д., ред. Желчные кислоты, об. 2. Нью-Йорк: Нью-Йорк. Пленум Пресс; 1973.
Google ученый
28.
Bracco U. Влияние структуры триглицеридов на всасывание жира. Am J Clin Nutr. 1994; 60: 1002С – 9С.
CAS
PubMed
Google ученый
29.
Westergaard H, Dietshcy JM. Механизм, посредством которого мицеллы желчных кислот увеличивают скорость поглощения жирных кислот и холестерина клетками слизистой оболочки кишечника. J Clin Invest. 1976; 58: 97–108.
CAS
PubMed Central
PubMed
Google ученый
30.
Джонстон Дж. М., Боргстром Б. Кишечная абсорбция и метаболизм мицеллярного раствора на липидах. Biochem Biophys Acta. 1964; 84: 412–23.
CAS
PubMed
Google ученый
31.
Chow SL, Hollander D. Двойной, зависящий от концентрации механизм абсорбции линолевой кислоты тонкой кишкой крысы in vitro. J Lipid Res. 1979; 20: 349–56.
CAS
PubMed
Google ученый
32.
Киндель Т., Ли Д.М., Цо П. Механизм образования и секреции хиломикронов. Atheroscler Suppl. 2010; 11: 11–6.
CAS
PubMed
Google ученый
33.
Stremmel W, Pohl L, Ring A, Hermann T. Новая концепция клеточного поглощения и внутриклеточного переноса длинноцепочечных жирных кислот. Липиды. 2001; 36: 981–9.
CAS
PubMed
Google ученый
34.
Мартинес-Августин О., Санчес де Медина Ф. Физиология и патофизиология кишечных желчных кислот. Мир Дж. Гастроэнтерол. 2008; 14: 5630–40.
CAS
PubMed Central
PubMed
Google ученый
35.
Cunningham HM, Leat WMF. Синтез липидов моноглицеридным и α-глицерофосфатным путями в кишечнике овец. Может J Biochem. 1969; 47: 1013–20.
CAS
PubMed
Google ученый
36.
Sabesin SM, Frase S. Электронно-микроскопические исследования сборки, внутриклеточного транспорта и секреции хиломикронов кишечником крысы. J Lipid Res. 1977; 18: 496–511.
CAS
PubMed
Google ученый
37.
Wang H, Eckel RH. Липопротиен липаза: от гена к ожирению. Обзор Am J Physiol Endocrinol Metab A. 2009; 297: 271–88.
Google ученый
38.
Бабаян В.К. Триглицериды со средней длиной цепи и структурированные липиды. Липиды. 1987; 22: 417–20.
CAS
PubMed
Google ученый
39.
Phan CT, Tso P. Абсорбция и транспорт липидов в кишечнике. Передние биоски.2001; 6: D299–319.
Азаин МДж. Жир в корме свиней. В: Льюис AJ, Southern LL, редакторы. Питание свиней. Бока-Ратон: CRC Press; 2001. с. 95–106.
Google ученый
42.
Lin X, Azain M, Odle J. Липиды и утилизация липидов у свиней. В: Л.И. Чиба, редактор, Sustainable Swine Nutrition . Blackwell Publishing Ltd., Оксфорд, Великобритания. 2013, стр. 59-79
43.
Бабатунде GM, Pond WG, Walker Jr EF, Chapman P, Banis RJ. Гематологические изменения, изменения кожи и очевидная усвояемость липидов и белка у растущих свиней мужского и женского пола, которых кормили рационами, содержащими сафлоровое масло, гидрогенизированное кокосовое масло, холестерин или вообще без жира. J Anim Sci. 1968; 27: 985–91.
Google ученый
44.
Cera KR, Mahan DC, Reinhart GA. Еженедельная усвояемость рациона свиней-отъемышей с добавлением кукурузного масла, сала или сала. J Anim Sci. 1988; 66: 1430–7.
CAS
PubMed
Google ученый
45.
Cera KR, Mahan DC, Reinhart GA. Влияние диетической сухой сыворотки и кукурузного масла на продуктивность поросят-отъемышей, усвояемость жира и использование азота.J Anim Sci. 1988; 666: 1438–45.
Google ученый
46.
Cera KR, Mahan DC, Reinhart GA. Очевидная перевариваемость жиров и показатели эффективности рационов для свиней после отъема, содержащих кокосовое масло, кукурузное масло или жир. J Anim Sci. 1989; 67: 2040–7.
Google ученый
47.
Cera KR, Mahan DC, Reinhart GA. Оценка различных экстрагированных растительных масел, жареных соевых бобов, триглицеридов со средней длиной цепи и смеси животных и растительных жиров для свиней после отъема.J Anim Sci. 1990; 68: 2756–65.
CAS
PubMed
Google ученый
48.
Li DF, Thaler RC, Nelssen JL, Harmon DL, Allee GL, Weeden TL. Влияние источников и комбинаций жиров на продуктивность стартовых свиней, усвояемость питательных веществ и морфологию кишечника. J Anim Sci. 1990; 68: 3694–704.
CAS
PubMed
Google ученый
49.
Джонс Д.Д., Хэнкок Д.Д., Хармон Д.Л., Уокер К.Э.Влияние экзогенных эмульгаторов и источников жира на усвояемость питательных веществ, липиды сыворотки и показатели роста поросят-отъемышей. J Anim Sci. 1992; 70: 3473–82.
CAS
PubMed
Google ученый
50.
Йоргенсен Х., Габерт В.М., Хедеманн М.С., Йенсен СК. Переваривание жира не отличается у растущих свиней, которых кормят рационами, содержащими рыбий жир, рапсовое масло или кокосовое масло. J Nutr. 2000; 130: 852–7.
CAS
PubMed
Google ученый
51.
Lowrey RS, Pond WG, Loosli JK, Maner JH. Влияние уровня пищевых жиров на очевидную усвояемость питательных веществ при выращивании свиней. J Anim Sci. 1962; 21: 746–50.
Google ученый
52.
Cera KR, Mahan DC, Reinhart GA. Показатели продуктивности свиней после отъема и реакции сывороточного профиля на дополнительные среднецепочечные свободные жирные кислоты и жир. J Anim Sci. 1989; 67: 2048–55.
CAS
Google ученый
53.
Li S, Sauer WC. Влияние содержания жира в рационе на усвояемость аминокислот у молодых свиней. J Anim Sci. 1994; 72: 1737–43.
CAS
PubMed
Google ученый
54.
Jorgensen H, Fernandez JA. Химический состав и энергетическая ценность различных источников жира для выращивания свиней. Acta Agric Scand Sect A Animal Sci. 2000. 50: 129–36.
CAS
Google ученый
55.
Гамильтон RMG, McDonald BE. Влияние источника диетического жира на очевидную перевариваемость жира и состав фекальных липидов молодой свиньи. J Nutr. 1969; 97: 33–41.
CAS
PubMed
Google ученый
56.
Frobish LT, Hays VW, Speer VC, Ewan RC. Влияние источника и уровня жира на использование жира молодыми свиньями. J Anim Sci. 1970; 30: 197–202.
CAS
PubMed
Google ученый
57.
Паулс Дж., Вайзман Дж., Коул Д. А., Джаггер С. Прогноз очевидной усвояемой энергетической ценности жиров, даваемых свиньям. Anim Sci. 1995; 61: 149–54.
Google ученый
58.
Уайзман Дж., Поулз Дж., Сальвадор Ф. Сравнение свиней и домашней птицы в прогнозировании пищевой энергетической ценности жиров. Anim Feed Sci Tech. 1998; 71: 1–9.
Google ученый
59.
Brambila S, Hill FW.Сравнение нейтрального жира и свободных жирных кислот в диетах с высоким содержанием липидов и низким содержанием углеводов для растущей курицы. J Nutr. 1966; 88: 84–92.
CAS
PubMed
Google ученый
60.
Mendoza SM, van Heugten E. Влияние источников липидов в рационе на продуктивность и очевидную общую усвояемость липидов и энергии в пищеварительном тракте при кормлении свиноматками. J Anim Sci. 2014; 92: 627–36.
CAS
PubMed
Google ученый
61.
Rosero DS, Odle J, Arellano C, Boyd RD, van Heugten E. Разработка прогнозных уравнений для оценки кажущейся энергии усвояемости липидов при скармливании лактирующим свиноматкам. J Anim Sci. 2015; 93: 1165–76.
CAS
PubMed
Google ученый
62.
Wiseman J, Salvador F. Влияние содержания свободных жирных кислот и степени насыщения на кажущуюся метаболизируемую энергетическую ценность жира, скармливаемого бройлерам. Poult Sci.1991; 70: 573–82.
CAS
PubMed
Google ученый
63.
Паулс Дж., Уайзман Дж., Коул Д. Д. А., Харди Б. Влияние химической структуры жиров на их кажущуюся усвояемую энергетическую ценность при введении молодым свиньям. Anim Prod. 1994; 58: 411–7.
CAS
Google ученый
64.
DeRouchey JM, Hancock JD, Hines RD, Maloney CA, Lee DJ, Cao H, et al. Влияние прогорклости и свободных жирных кислот в отборном белом жире на показатели роста и усвояемость питательных веществ у поросят-отъемышей.J Anim Sci. 2004. 82: 2937–44.
CAS
PubMed
Google ученый
65.
Керр Б.Дж., Шурсон Г.К. Перевариваемость липидов и энергетическая ценность кукурузного и соевого масла, содержащего различные уровни свободных жирных кислот, скармливаемых родительским свиньям. J Anim Sci 2015, 93 (Дополнение 1): В ПРЕССЕ.
66.
Таллис Дж. Б., Виттемор, Коннектикут. Усвояемость полностью гидрогенизированного жира для выращивания свиней. Anim Feed Sci Tech. 1980; 5: 87–91.
CAS
Google ученый
67.
Гатлин Л.А., см. MT, Одле Дж. Влияние химического гидрирования дополнительного жира на относительную очевидную перевариваемость липидов у откормочных свиней. J Anim Sci. 2005; 83: 1890–8.
CAS
PubMed
Google ученый
68.
Оверленд М., Токач М.Д., Корнелиус С.Г., Петтигрю Дж. Э., Руст Дж. У. Лецитин в рационе свиней: I. Поросята-отъемыши. J Anim Sci. 1993; 71: 1187–93.
CAS
PubMed
Google ученый
69.
Overland M, Tokach MD, Cornelius SG, Pettigrew JE, Wilson MW. Лецитин в рационах свиней: II Свиньи на откорме. J Anim Sci. 1993; 71: 1194–7.
CAS
PubMed
Google ученый
70.
Оверленд М., Мроз З., Сундстол Ф. Влияние лецитина на кажущуюся подвздошную и общую усвояемость сырого жира и жирных кислот у свиней. J Anim Sci. 1994; 72: 2022–8.
CAS
PubMed
Google ученый
71.
deSouza TR, Peiniau J, Mounier A, Aumaitre A. Влияние добавления жира и лецитина в рацион поросят-отъемышей на видимый общий тракт и перевариваемость жиров и жирных кислот в подвздошной кишке. Anim Feed Sci Tech. 1995; 52: 77–91.
Xing JJ, van Heugten E, Li DF, Touchette KJ, Coalson JA, Odgaard RL и др. Влияние эмульгирования, инкапсуляции жира и гранулирования на продуктивность поросят-отъемышей и усвояемость питательных веществ. J Anim Sci. 2004; 82: 2601–9.
CAS
PubMed
Google ученый
74.
Реннер Р., Хилл ФВ. Факторы, влияющие на всасываемость насыщенных жирных кислот цыпленком. J Nutr. 1961; 74: 254–8.
CAS
Google ученый
75.
Brink EJ, Haddeman E, de Fouw JJ, Weststrate JA. Позиционное распределение стеариновой кислоты и олеиновой кислоты в триацилглицерине и концентрация кальция в пище определяют кажущееся всасывание этих жирных кислот у крыс. J Nutr. 1995; 125: 2379–87.
CAS
PubMed
Google ученый
76.
Брокерхофф Х. Стереоспецифический анализ триглицеридов. Липиды. 1971; 4: 942–56.
Google ученый
77.
Smink W, Gerrits WJJ, Hovenier R, Geelen MJH, Lobee HWJ, Verstegan MWA и др. Переваривание и отложение жирных кислот в рационах цыплят-бройлеров, содержащих либо натуральное, либо рандомизированное пальмовое масло. Poult Sci. 2008; 87: 506–13.
CAS
PubMed
Google ученый
78.
Scheeder MRL, Gumy D, Messikommer R, Wenk C., Lambelet P. Влияние ПНЖК в положении sn-2 в диетических триацилглицеринах на состав жирных кислот жировой ткани нежвачных сельскохозяйственных животных.Eur J Lipid Sci Technol. 2003. 105: 74–82.
CAS
Google ученый
79.
Innis SM, Dyer R, Quinlan PT, Diersen-Schade D. Диетическая структура триацилглицерина и насыщенные жиры изменяют жирные кислоты плазмы и тканей у поросят. Липиды. 1996; 31: 497–505.
CAS
PubMed
Google ученый
80.
Innis SM, Dyer R. Диетические триацилглицерины с пальмитиновой кислотой (16: 0) во 2-м положении увеличиваются на 16: 0 во 2-м положении триацилглицеринов плазмы и хиломикрон, но снижают фосфолипид арахидоновую и докозагексаеновую кислоты, и изменяют метаболизм сложного холестерилового эфира у поросят, получающих смесь.J Nutr. 1997; 127: 1311–9.
CAS
PubMed
Google ученый
81.
Гриффит. Влияние пищевых жиров и целлюлозы на очевидную усвояемость кальция у растущих цыплят. Poult Sci. 1961; 40: 1492–7.
CAS
Google ученый
82.
Аттех Дж.О., Лисон С. Влияние возраста, содержания холевой кислоты и кальция в пище на продуктивность, использование свободных жирных кислот и минерализацию костей у бройлеров.Poult Sci. 1985; 64: 1959–71.
CAS
PubMed
Google ученый
83.
Wiseman J, Cole DJA, Hardy B. Пищевая энергетическая ценность соевого масла, жира и их смесей для выращивания / откорма свиней. Anim Prod. 1990; 50: 513–8.
Google ученый
84.
Паулс Дж., Уайзман Дж., Коул Д. Д. А., Харди Б. Влияние химической структуры жиров на их кажущуюся усвояемую энергетическую ценность при употреблении растущих / откормочных свиней.Anim Prod. 1993; 57: 137–46.
CAS
Google ученый
85.
Керр Б.Дж., Вебер Т.Э., Дозье III ВА, Кидд М.Т. Содержание усвояемой и метаболизируемой энергии сырого глицерина, происходящего из различных источников, у поросят. J Anim Sci. 2009; 87: 4042–9.
CAS
PubMed
Google ученый
86.
Silva HO, Sousa RV, Fialho ET, Lima JAF, Silva LF. Усвояемая и метаболическая энергия масел и сала для выращивания свиней.J Anim Sci. 2009; 87 (Е-Дополнение 2): 63. Abstr.
Google ученый
87.
Андерсон П.В., Керр Б.Дж., Вебер Т.Е., Цимер С.Дж., Шурсон Г.К. Определение и прогнозирование энергии на основе химического анализа побочных продуктов кукурузы, поступающих на откорма свиней. J Anim Sci. 2012; 90: 1242–54.
CAS
PubMed
Google ученый
88.
NRC. Потребности свиней в питательных веществах. 10 об. Изд. Вашингтон, округ Колумбия: Natl.Акад. Нажмите; 1998.
Google ученый
89.
Ле Гофф Дж., Ноблет Дж. Сравнительная перевариваемость пищевой энергии и питательных веществ в тракте у растущих и взрослых свиноматок. J Anim Sci. 2001; 79: 2418–27.
PubMed
Google ученый
90.
Штайн Х. Х., Ким С.В., Нильсен ТТ, Пасха, РА. Стандартизированная усвояемость белков подвздошной кишки и аминокислот растущими свиньями и свиноматками. J Anim Sci.2001; 79: 2113–22.
CAS
PubMed
Google ученый
91.
Кил Д.Й., Джи Ф., Стюарт Л.Л., Хинсон Р.Б., Болье А.Д., Аллее Г.Л. и др. Чистая энергия соевого масла и отборного белого жира в рационах для свиней на доращивании и откорме. J Anim Sci. 2011; 89: 448–59.
CAS
PubMed
Google ученый
92.
ван Милген Дж, Ноблет Дж, Дубиос С. Энергетическая эффективность использования крахмала, белка и липидов у растущих свиней.J Nutr. 2001; 131: 1309–18.
PubMed
Google ученый
93.
Just A. Чистая энергетическая ценность сырого жира для роста свиней. Livest Prod Sci. 1982; 9: 501–9.
Google ученый
94.
Ноблет Дж, Форчун Х, Дюпир С., Дюбуа С. Усвояемые, метаболизируемые и полезные значения энергии 13 кормов для растущих свиней: Влияние энергетической системы. Anim Feed Sci Tech. 1993; 42: 131–49.
CAS
Google ученый
95.
Halas VL, Babinszky L, Dijkstra J, Verstegen MWA, Gerrits WJJ. Эффективность отложения жира из некрахмальных полисахаридов, крахмала и ненасыщенных жиров у свиней. Br Jour Nutr. 2010; 97: 33–41.
Google ученый
96.
Sauvant D, Perex JM, Tran G. Таблицы состава и пищевой ценности кормовых материалов, INRA, Париж, Франция.Вагенинген, Нидерланды: Академические издательства Вагенингена; 2004.
Google ученый
97.
Galloway ST, Ewan RC. Энергетическая оценка сала и овсяной крупы для молодняка свиней. J Anim Sci. 1989; 67: 1744–50.
Google ученый
98.
Ewan RC. Прогнозирование использования энергии рационами и ингредиентами кормов свиньями. В: van det Honing Y, Close WH, editors. Энергетический метаболизм, бюллетень Европейской ассоциации животноводства No.43. Пудок Вагенинген, Нидерланды. 1989. стр. 271–4.
Schaich KM. Окисление липидов: теоретические аспекты. В: Промышленные масложировые продукты Bailey’s, Vol. 1. Пищевые масла и жировые продукты: химия, свойства и влияние на здоровье. Хобокен, штат Нью-Джерси: «Джон Уайли и сыновья, Inc.»; 2005 г.п. 269–355.
Google ученый
103.
Лабуза ТП. Кинетика окисления липидов в пищевых продуктах. В CRC Critical Rev Food Tech. 1971; 2: 355–405.
Google ученый
104.
Gutteridge JMC. Перекисное окисление липидов и антиоксиданты как биомаркеры повреждения тканей. Clin Chem. 1995; 41: 1819–28.
CAS
PubMed
Google ученый
105.
Сант Анджело AJ. Окисление липидов в пищевых продуктах. Crit Rev Food Sci Nutr. 1996; 36: 175–224.
CAS
PubMed
Google ученый
106.
Навар WW. Липиды, гл. 5, в Food Chemistry, 3 -е изд. , редактор О. Р. Феннема. Марсель Деккер, Инк., Нью-Йорк, штат Нью-Йорк. 1996, стр. 225-319
107.
Schaich KM. Мышление за пределами классической цепной реакции окисления липидов. Lipid Tech. 2012; 24: 55–8.
CAS
Google ученый
108.
Холман РТ. Автоокисление жиров и родственных веществ. В: Холман Р.Т., Лундберг В.О., Малкин Т., редакторы. Прогресс в химии жиров и других липидов. Лондон: Pergamon Press; 1954. с. 51–98.
Google ученый
109.
Науди А., Джов М., Аяла В., Рамирес О., Кабре Р., Прат Дж. И др. Регионоспецифическая уязвимость центральной нервной системы человека к перекисному окислению липидов. В: Перекисное окисление липидов А. Катала, редактор. Intech. 2012 г.п. 437–56.
Google ученый
110.
Lau FY, Hammond EG, Ross PF. Влияние рандомизации на окисление кукурузного масла. JAOCS. 1982; 59: 407–11.
CAS
Google ученый
111.
Tautorus CL, McCurdy AR. Влияние рандомизации на окислительную стабильность растительных масел при двух различных температурах. JAOCS. 1990; 67: 525–30.
CAS
Google ученый
112.
Tautorus CL, McCurdy AR. Влияние рандомизации на стабильность смесей триолеилглицерина и льняного масла. JAOCS. 1992; 69: 538–44.
Ван Т., Цзян Дж., Хаммонд Э.Г.Влияние рандомизации на окислительную стабильность кукурузного масла. JAOCS. 2005; 82: 111–7.
CAS
Google ученый
115.
Choe E, Min DB. Химия фритюрных масел. J Food Sci. 2007; 72: R77–86.
CAS
PubMed
Google ученый
116.
Янеро ДР. Реактивность малонового диальдегида и тиобарбитуровой кислоты как диагностические показатели перекисного окисления липидов и перекисного повреждения тканей.Free Radic Biol Med. 1990; 9: 515–40.
CAS
PubMed
Google ученый
117.
Дель Рио Д., Стюарт А. Дж., Пеллегрини Н. Обзор недавних исследований малонового диальдегида как токсичной молекулы и биологического маркера окислительного стресса. Нутр Метаб Кардиоваск Дис. 2005; 15: 316–28.
PubMed
Google ученый
118.
Esterbauer H, Schaur RJ, Zollner H. Химия и биохимия 4-гидроксиноненаля, малонового альдегида и родственных альдегидов.Free Radic Biol Med. 1991; 11: 81–128.
CAS
PubMed
Google ученый
119.
Poli G, Schaur RJ, Siems WG, Leonarduzzi G. 4-Hydroxynonenal: продукт окисления мембранных липидов, представляющий медицинский интерес. Med Res Rev.2008; 28: 569–631.
CAS
PubMed
Google ученый
120.
Ники Э. Перекисное окисление липидов: физиологические уровни и двойные биологические эффекты.Free Radic Biol Med. 2009; 47: 469–84.
CAS
PubMed
Google ученый
121.
Спителлер Г. Пероксирадикалы: индукторы нейродегенеративных и других воспалительных заболеваний. Их происхождение и то, как они превращают холестерин, фосфолипиды, плазмалогены, полиненасыщенные жирные кислоты, сахара и белки в вредные продукты. Free Radic Biol Med. 2006; 41: 362–87.
CAS
PubMed
Google ученый
122.
Marquez-Ruiz G, Perez-Camino MC, Dobarganes MC. Комбинация адсорбционной и эксклюзионной хроматографии для определения мономеров, димеров и полимеров жирных кислот. J Хромат. 1990; 514: 37–44.
CAS
Google ученый
123.
ИЮПАК. Стандартный метод 2.508: Определение полимеризованных триглицеридов в маслах и жирах с помощью высокоэффективной жидкостной хроматографии. В кн .: Стандартные методы анализа масел, жиров и их производных.7-е изд. Блэквелл, Оксфорд: Международный союз теоретической и прикладной химии; 1992.
Google ученый
124.
Ким Р.С., ЛаБелла Ф.С. Сравнение аналитических методов мониторинга профилей автоокисления аутентичных липидов. J Lipid Res. 1987. 28: 1110–7.
CAS
PubMed
Google ученый
125.
Шахиди Р., Чжун Ю. Окисление липидов: методы измерения.В: Промышленные масложировые продукты Bailey’s, Vol. 1. Пищевые масла и жировые продукты: химия, свойства и влияние на здоровье. Хобокен, Нью-Джерси: Джон Уайли и сыновья; 2005. с. 357–85.
Google ученый
126.
Шахиди Р., Ванасундара Дж. Методы оценки окислительной стабильности липидсодержащих пищевых продуктов. Food Sci Technol Int. 1996; 2: 73–81.
CAS
Google ученый
127.
Лю П., Керр Б.Дж., Чен С., Вебер Т.Е., Джонстон Л.Дж., Шурсон Г.К. Методы создания термически окисленных липидов и сравнение аналитических процедур для характеристики перекисного окисления. J Anim Sci. 2014; 92: 2950–9.
CAS
PubMed
Google ученый
128.
ван Кемпен Т.А., МакКомас С. Инфракрасная спектроскопия как инструмент оценки качества жира. J Appl Poult Res. 2002; 11: 191–201.
Google ученый
129.
Такахаши К., Акиба Ю. Влияние окисленного жира на продуктивность и некоторые физиологические реакции у цыплят-бройлеров. Jap Poult Sci. 1999; 36: 304–10.
CAS
Google ученый
130.
Болер Д., Фернандес-Дуэньяс Д., Кутцлер Л., Чжао Дж., Харрелл Р., Кэмпион Д. и др. Влияние окисленного кукурузного масла и смеси синтетических антиоксидантов на продуктивность, окислительный статус тканей и качество свежего мяса в откормочных курганах. J Anim Sci.2012; 90: 5159–69.
CAS
PubMed
Google ученый
131.
Лю П., Чен С., Керр Б.Дж., Вебер Т.Е., Джонстон Л.Дж., Шурсон Г.К. Влияние термически окисленных растительных масел и животных жиров на энергию и усвояемость питательных веществ у молодняка свиней. J Anim Sci. 2014; 92: 2971–9.
CAS
PubMed
Google ученый
132.
Betteridge DJ. Что такое оксидативный стресс? Обмен веществ.2000; 49: 3–8.
CAS
PubMed
Google ученый
133.
Ликкесфельдт Дж., Свендсен О. Окислители и антиоксиданты при болезни: окислительный стресс у сельскохозяйственных животных. Вет Дж. 2007; 173: 502–11.
Роби В., Шермер В. Повреждающее действие окисления. Кормовая смесь. 1994; 2: 22–6.
Google ученый
136.
Дибнер Дж., Васкес-Анон М., Найт С. Понимание окислительного баланса и его влияния на продуктивность животных. В: Proceedings 2011 Корнельская конференция по питанию для производителей кормов, Восточные Сиракузы, Нью-Йорк. 2011. с. 1–7.
Google ученый
137.
МакГилл Дж., МакГилл Э., Камьяб А., Фирман Дж.Влияние жиров с высоким перекисным числом на продуктивность бройлеров с нормальным иммунным состоянием. Int J Poult Sci. 2011; 10: 241–6.
CAS
Google ученый
138.
МакГилл Дж., МакГилл Е., Камьяб А., Фирман Дж., Руис-Фериа С., Ларрисон Е. и др. Влияние жиров с высоким перекисным числом на продуктивность бройлеров в состоянии иммунной защиты. Int J Poult Sci. 2011; 10: 665–9.
CAS
Google ученый
139.
Таварес М.А., Болер Д.Д., Бесс К.Н., Чжао Дж., Ян Ф., Дилгер А.С. и др. Влияние включения антиоксидантов и качества масла на продуктивность бройлеров, качество мяса и окисление липидов. Poult Sci. 2011; 90: 922–30.
PubMed
Google ученый
140.
Лю П., Чен С., Керр Б.Дж., Вебер Т.Е., Джонстон Л.Дж., Шурсон Г.К. Влияние термически окисленных растительных масел и животных жиров на показатели роста, экспрессию генов печени, холестерин и триглицериды в печени и сыворотке молодых свиней.J Anim Sci. 2014; 92: 2960–70.
CAS
PubMed
Google ученый
141.
Иноуэ Т., Курашиге А., Минетома Т., Шигё Ф. Питательный эффект окисленного соевого масла в рационе бройлеров. В: Материалы XVII Всемирного птицеводческого конгресса, Хельсинки, Финляндия. 1984. с. 368–9.
Google ученый
142.
Энгберг Р.М., Лауридсен С., Йенсен С.К., Якобсен К. Включение окисленного растительного масла в рационы бройлеров.Его влияние на баланс питательных веществ и антиоксидантный статус бройлеров. Poult Sci. 1996; 75: 1003–11.
CAS
PubMed
Google ученый
143.
Анджум М., Мирза И., Хан А., Азим А. Влияние свежего и окисленного соевого масла на показатели роста, вес органов и качество мяса цыплят-бройлеров. Пакистан Вет Дж. 2004; 24: 173–8.
CAS
Google ученый
144.
Дибнер Дж., Китчелл М., Этвелл С., Айви Ф. Влияние пищевых ингредиентов и возраста на микроскопическую структуру желудочно-кишечного тракта у домашней птицы. J Appl Poultry Res. 1996; 5: 70–7.
Google ученый
145.
Asghar A, Lin C, Gray J, Buckley D, Booren A, Crackel R, et al. Влияние окисленных диетических масел и добавок антиоксидантов на стабильность мембраносвязанных липидов в мясе бройлеров. Br Poult Sci. 1989; 30: 815–23.
CAS
PubMed
Google ученый
146.
Racanicci AMC, Menten JFM, Regitano-d’Arce MAB, Torres EAFS, Pino LM, Pedroso AA. Окисленные пищевые жирные субпродукты птицы: продуктивность бройлеров и устойчивость мяса бедра к окислению при хранении в охлажденном виде. Rev Bras Cienc Aví. 2008; 10: 29–35.
Google ученый
147.
Холливелл B, Мурсия, Массачусетс, Кирико S, Аруома О.И.Свободные радикалы и антиоксиданты в продуктах питания и in vivo: что они делают и как работают. Crit Rev Food Sci Nutr. 1995; 35: 7–20.
CAS
PubMed
Google ученый
148.
Reichling JJ, Kaplan MM. Клиническое применение сывороточных ферментов при заболеваниях печени. Dig Dis Sci. 1988; 33: 1601–14.
CAS
PubMed
Google ученый
149.
Тейге Дж., Нордстога К., Аурсьо Дж. Влияние диеты на экспериментальную дизентерию свиней.1. Эффекты диеты с дефицитом витамина Е и селена с добавлением 6,8% жира печени трески. Acta Vet Scand. 1977; 18: 384–96.
CAS
PubMed
Google ученый
150.
van Vleet JF. Сравнительная эффективность пяти процедур приема добавок для контроля дефицита селена и витамина Е у свиней. Am J Vet Res. 1982; 43: 1180–9.
PubMed
Google ученый
151.
Хоссейн Садрзаде С., Нанджи А.А., Мейдани М. Влияние хронического кормления этанолом на уровни α- и γ-токоферола в плазме и печени у нормальных крыс и крыс с дефицитом витамина E: взаимосвязь с перекисным окислением липидов. Biochem Pharmacol. 1994; 47: 2005–10.
Google ученый
152.
Лю Дж.Ф., Хуанг СиДжей. Удержание тканевого альфа-токоферола у самцов крыс нарушается при кормлении рационами, содержащими окисленное масло для жарки. J Nutr. 1995; 125: 3071–9.
CAS
PubMed
Google ученый
153.
Яник Ф, Аманвермез Р., Яник А., Челик С., Кёкчу А. Преэклампсия и эклампсия, связанные с повышенным перекисным окислением липидов и снижением уровня витамина Е в сыворотке. Int J Gynecol Obstet. 1999. 64: 27–33.
CAS
Google ученый
154.
Джонс Д.П. Редокс-потенциал пары GSH / GSSG: анализ и биологическое значение. Методы Энзимол. 2002; 348: 93–112.
CAS
PubMed
Google ученый
155.
Антолович М., Пренцлер П.Д., Пацалидес Э., Макдональд С., Робардс К. Методы тестирования антиоксидантной активности. Аналитик. 2002; 127: 183–98.
CAS
PubMed
Google ученый
156.
Sies H. Общая антиоксидантная способность: оценка концепции. J Nutr. 2007; 137: 1493–5.
CAS
PubMed
Google ученый
157.
Шервин Э. Окисление и антиоксиданты при переработке масел и жиров.J Am Oil Chem Soc. 1978; 55: 809–14.
CAS
Google ученый
158.
Ванасундара PKJPD, Шахиди Ф. Антиоксиданты: наука, технология и приложения. Гл. 11, в Bailey’s Industrial Oil and Fat Products, 6-е изд. John Wiley & Sons, Inc., Хобокен, Нью-Джерси. 2005, стр. 431-489.
Gordon MH. Механизм антиоксидантного действия in vitro. В: Hudson BJF, редактор. Пищевые антиоксиданты. Нью-Йорк: Спрингер; 1990. стр. 1–18.
Google ученый
161.
Франкель Э., Куни П., Мозер Х, Коуэн Дж., Эванс С. Влияние антиоксидантов и инактиваторов металлов в соевом масле без токоферола. Fette Wiss Technol. 1959; 61: 1036–9.
CAS
Google ученый
162.
Флидер Ф., Ортофер Ф. Металлы в соевом масле. J Am Oil Chem Soc. 1981; 58: 270–2.
CAS
Google ученый
163.
Клементс А., Ван Ден Энг Р., Фрост Д., Хугенхаут К., Нуй Дж. Участие синглетного кислорода в фотосенсибилизированном окислении 1,4-диеновых систем и фотоокислении соевого масла. J Am Oil Chem Soc. 1973; 50: 325–30.
CAS
PubMed
Google ученый
164.
Cort WM. Антиоксидантные свойства аскорбиновой кислоты в пищевых продуктах. В: Сейб П., редактор. Успехи в серии химии. Вашингтон, округ Колумбия: Американское химическое общество; 1982. с. 533–50.
Google ученый
165.
Фернандес-Дуэньяс DM. Влияние окисленного кукурузного масла и синтетического антиоксиданта на продуктивность свиней, антиоксидантный статус тканей, качество свинины и оценка срока хранения. В: к.т.н. диссертация, Урбана, Иллинойс. 2009.
Google ученый
166.
Дибнер Дж., Этвелл С., Китчелл М., Шермер В., Айви Ф. Кормление бройлеров и свиней окисленными жирами: влияние на обмен энтероцитов, пролиферацию гепатоцитов и лимфоидную ткань, связанную с кишечником. Anim Feed Sci Technol. 1996; 62: 1–13.
CAS
Google ученый
167.
Harrell RJ, Zhao J, Reznik G, Macaraeg D, Wineman T, Richards J. Применение смеси диетических антиоксидантов у поросят, которых кормили свежим или окисленным кукурузным маслом DDGS.J Anim Sci. 2010; 88 (Е-Дополнение 3): 60. Abstr.
Google ученый
168.
Лу Т., Харпер А.Ф., Чжао Дж., Эстьен М.Дж., Даллул, РА. Добавление антиоксидантов в рацион свиней с высоким содержанием оксидантов: I. Влияние на показатели роста, функцию печени и окислительный статус. J Anim Sci. 2014; 92: 5455–63.
CAS
PubMed
Google ученый
169.
Ван С.Ю., Боттье В., Мейнард П., Дибнер Дж., Шермер В.Влияние сантохина и окисленного жира на глутатион в печени и кишечнике у бройлеров. Poult Sci. 1997. 76: 961–7.
CAS
PubMed
Google ученый
170.
Анджум Мичиган, Алам МЗ, Мирга И. Влияние неокисленного и окисленного соевого масла с добавлением двух уровней антиоксиданта на продуктивность бройлеров. Азиатско-Aust J Anim Sci. 2002; 15: 713–20.
CAS
Google ученый
171.
Fernández-Dueñas DM, Mariscal G, Ramírez E, Cuarón JA. Витамин С и β-каротин в рационах свиней при отъеме. Anim Feed Sci Technol. 2008; 146: 313–26.
Google ученый
172.
Сонг Р., Чен С., Ван Л., Джонстон Л.Дж., Керр Б.Дж., Вебер Т.Э. и др. Высокое содержание серы в зерне сушеных дистилляторов кукурузы с растворимыми (DDGS) защищает от окисленных липидов в DDGS за счет увеличения содержания серосодержащих антиоксидантов у поросят. J Anim Sci.2013; 91: 2715–28.
CAS
PubMed
Google ученый
173.
Сонг Р., Чен С., Джонстон Л.Дж., Керр Б.Дж., Вебер Т.Э., Шурсон Г.К. Влияние рационов кормления, содержащих высокопероксидные дистилляторы, сушеные зерна с растворимыми веществами и повышение уровня витамина Е к концу отъема, на показатели роста, характеристики туши и состав свиного жира. J Anim Sci. 2014; 92: 198–210.
CAS
PubMed
Google ученый
174.
USDA. База данных Министерства сельского хозяйства США по способности поглощать радикалы кислорода (ORAC) отобранных пищевых продуктов, выпуск 2. Белтсвилл, Мэриленд: Служба сельскохозяйственных исследований Министерства сельского хозяйства США; 2010. с. 1–48.
Пакер Л., Вебер С.У., Римбах Г. Молекулярные аспекты антиоксидантного действия α-токотриенола и клеточная передача сигналов.J Nutr. 2001; 131: 369С – 73С.
CAS
PubMed
Google ученый
177.
Lauridsen C, Engel H, Craig AM, Traber M. Относительная биоактивность диетических RRR- и all-rac-альфа-токоферилацетатов у свиней, оцененная с помощью меченного дейтерием витамина E. J Anim Sci. 2002; 80: 702–7.
CAS
PubMed
Google ученый
178.
Лауридсен К., Энгель Х., Йенсен С.К., Крейг А.М., Трабер М.Г.Кормящие свиноматки и поросята-сосуны предпочтительно включают RRR-альфа-токоферол в молоко, плазму и ткани. J Nutr. 2002; 132: 1258–64.
CAS
PubMed
Google ученый
179.
Podda M, Weber C, Traber MG, Packer L. Одновременное определение тканевых токоферолов, токотриенолов, убихинолов и убихинонов. J Lip Res. 1996; 37: 893–901.
CAS
Google ученый
180.
Ullrey DE. Витамин Е для свиней. J Anim Sci. 1981; 53: 1039–56.
CAS
PubMed
Google ученый
181.
Чанг Й, Махан Д., Лепин А. Эффективность диетического d-альфа-токоферола и dl-альфа-токоферилацетата для поросят-отъемышей. J Anim Sci. 1992; 70: 2485–92.
CAS
PubMed
Google ученый
182.
Gropper SS, Smith JL. Жирорастворимые витамины.В: Гроппер С.С., Смит Дж. Л., редакторы. Продвинутое питание и метаболизм человека. Индепенденс, штат Кентукки: Обучение в Уодсворт Cengage; 2009. с. 371–424.
Google ученый
183.
Блохина О., Виролайнен Э., Фагерстедт К.В. Антиоксиданты, окислительное повреждение и кислородный стресс: обзор. Энн Бот. 2003. 91: 179–94.
CAS
PubMed Central
PubMed
Google ученый
184.
Sies H, Stahl W. Витамины E и C, бета-каротин и другие каротиноиды в качестве антиоксидантов. Am J Clin Nutr. 1995; 62: 1315С – 21С.
CAS
PubMed
Google ученый
185.
Ди Мацио П., Мерфи М.Э., Сайс Х. Системы антиоксидантной защиты: роль каротиноидов, токоферолов и тиолов. Am J Clin Nutr. 1991; 53: 194С – 200С.
Атмака Г. Антиоксидантные эффекты серосодержащих аминокислот. Йонсей Мед Дж. 2004; 45: 776–88.
CAS
PubMed
Google ученый
188.
Скримджер и Харвуд. Жирные кислоты и липидный состав. В: Gunstone FD, Harwood JL, Dijkstra AJ, редакторы.Справочник по липидам. 3-е изд. 2007. с. 1–36.
Google ученый
189.
AOCS. Физико-химические характеристики масел, жиров и восков, 2 -е издание . Д. Файерстоун изд. AOCS Press. Урбана, Иллинойс, 2006
190.
Gunstone FD, Harwood JL. Возникновение и характеристика масел и жиров. В: Gunstone FD, Harwood JL, Dijkstra AJ, редакторы. Справочник по липидам. 3-е изд. 2007. с. 37–141.
Google ученый
4.2: Что такое липиды? — Медицина LibreTexts
Навыки для развития
Объясните роль липидов в общем состоянии здоровья.
Липиды — важные жиры, которые выполняют разные функции в организме человека. Распространенное заблуждение состоит в том, что жир просто полнеет. Однако, вероятно, именно из-за жира мы все здесь. На протяжении всей истории было много случаев, когда еды не хватало. Наша способность накапливать избыточную калорийную энергию в виде жира для использования в будущем позволила нам продолжать жить как биологический вид во время голода.Итак, нормальные жировые запасы — это сигнал о том, что обменные процессы идут эффективно и человек здоров.
Липиды — это семейство органических соединений, которые в основном нерастворимы в воде. Липиды, состоящие из жиров и масел, представляют собой молекулы, которые выделяют высокую энергию и имеют химический состав в основном из углерода, водорода и кислорода. Липиды выполняют три основные биологические функции в организме: они служат в качестве структурных компонентов клеточных мембран, действуют как хранилища энергии и действуют как важные сигнальные молекулы.
Три основных типа липидов — это триацилглицерины (также называемые триглицеридами), фосфолипиды и стерины. Триацилглицерины (также известные как триглицериды) составляют более 95 процентов липидов в рационе и обычно содержатся в жареной пище, растительном масле, сливочном масле, цельном молоке, сыре, сливочном сыре и некоторых видах мяса. Натуральные триацилглицерины содержатся во многих продуктах питания, включая авокадо, оливки, кукурузу и орехи. Мы обычно называем содержащиеся в пище триацилглицерины «жирами» и «маслами».«Жиры — это липиды, твердые при комнатной температуре, а масла — жидкие. Как и большинство жиров, триацилглицерины не растворяются в воде. Термины «жиры», «масла» и «триацилглицерины» носят произвольный характер и могут использоваться как взаимозаменяемые. В этой главе, когда мы используем слово жир, мы имеем в виду триацилглицерины.
Фосфолипиды составляют лишь около 2 процентов пищевых липидов. Они водорастворимы и содержатся как в растениях, так и в животных. Фосфолипиды имеют решающее значение для создания защитного барьера или мембраны вокруг клеток вашего тела.Фактически, фосфолипиды синтезируются в организме с образованием мембран клеток и органелл. В крови и биологических жидкостях фосфолипиды образуют структуры, в которых жир заключен и транспортируется по кровотоку.
Рисунок 4.2.1: Типы липидов
Стерины — наименее распространенный тип липидов. Холестерин, пожалуй, самый известный стерол. Хотя холестерин имеет печально известную репутацию, организм получает лишь небольшое количество холестерина с пищей — организм производит большую часть этого холестерина.Холестерин является важным компонентом клеточной мембраны и необходим для синтеза половых гормонов, витамина D и солей желчных кислот.
Позже в этой главе мы рассмотрим каждый из этих липидов более подробно и узнаем, как функционируют их различные структуры, поддерживая работу вашего тела.
Функции липидов в организме: запасание энергии
Избыточная энергия пищи, которую мы едим, переваривается и включается в жировую ткань или жировую ткань. Большая часть энергии, необходимой человеческому организму, обеспечивается углеводами и липидами.Как обсуждалось в главе 3 «Углеводы», глюкоза хранится в организме в виде гликогена. В то время как гликоген является готовым источником энергии, липиды в первую очередь служат в качестве энергетического резерва. Как вы помните, гликоген довольно объемный и содержит много воды, поэтому организм не может хранить слишком много воды надолго. В качестве альтернативы жиры плотно упакованы без воды и хранят гораздо большее количество энергии в ограниченном пространстве. Грамм жира плотно сконцентрирован с энергией — он содержит более чем в два раза больше энергии, чем грамм углеводов.Энергия необходима для того, чтобы приводить в действие мышцы для всей физической работы и игры, в которой участвует средний человек или ребенок. Например, накопленная в мышцах энергия подталкивает спортсмена по трассе, подстегивает ноги танцора, чтобы продемонстрировать последние модные шаги, и удерживает все движущиеся части тела работают без сбоев.
В отличие от других клеток организма, которые могут накапливать жир в ограниченных количествах, жировые клетки специализируются на хранении жира и могут увеличиваться в размерах почти до бесконечности. Избыток жировой ткани может вызвать чрезмерную нагрузку на организм и нанести вред вашему здоровью.Серьезным следствием избытка жира является накопление слишком большого количества холестерина в стенке артерий, что может утолщать стенки артерий и приводить к сердечно-сосудистым заболеваниям. Таким образом, хотя некоторые жировые отложения имеют решающее значение для нашего выживания и хорошего здоровья, в больших количествах они могут быть препятствием для поддержания хорошего здоровья.
Функции липидов в организме: регулирование и передача сигналов
Триацилглицерины контролируют внутренний климат тела, поддерживая постоянную температуру. Те, у кого недостаточно жира в организме, как правило, быстрее простужаются, часто утомляются и имеют пролежни на коже из-за дефицита жирных кислот.Триацилглицерины также помогают организму вырабатывать и регулировать гормоны. Например, жировая ткань выделяет гормон лептин, регулирующий аппетит. В репродуктивной системе жирные кислоты необходимы для правильного репродуктивного здоровья; женщины, которым не хватает необходимого количества, могут прекратить менструацию и стать бесплодными. Незаменимые жирные кислоты омега-3 и омега-6 помогают регулировать уровень холестерина и свертываемость крови, а также контролировать воспаление в суставах, тканях и кровотоке. Жиры также играют важную функциональную роль в поддержании передачи нервных импульсов, хранении памяти и структуре тканей.В частности, в головном мозге липиды определяют активность мозга по структуре и функциям. Они помогают формировать мембраны нервных клеток, изолируют нейроны и способствуют передаче электрических импульсов по всему мозгу.
Знаете ли вы, что до 30 процентов веса тела состоит из жировой ткани? Некоторые из них состоят из висцерального жира или жировой ткани, окружающей нежные органы.Жизненно важные органы, такие как сердце, почки и печень, защищены висцеральным жиром. Состав мозга на 60% состоит из жира, что демонстрирует важную структурную роль, которую жир выполняет в организме. Возможно, вы больше всего знакомы с подкожным жиром или подкожным жиром. Этот покрывающий слой ткани изолирует тело от экстремальных температур и помогает контролировать внутренний микроклимат. Он накрывает наши руки и ягодицы и предотвращает трение, так как эти области часто соприкасаются с твердыми поверхностями.Это также дает телу дополнительную подкладку, необходимую при занятиях физически сложными видами деятельности, такими как катание на коньках или роликовых коньках, верховая езда или сноуборд.
Функции липидов в организме: помощь пищеварению и повышение биодоступности
Диетические жиры, содержащиеся в продуктах, которые мы едим, расщепляются в нашей пищеварительной системе и начинают транспортировку ценных питательных микроэлементов. Благодаря переносу жирорастворимых питательных веществ через процесс пищеварения кишечное всасывание улучшается.Это улучшенное всасывание также известно как повышенная биодоступность. Жирорастворимые питательные вещества особенно важны для хорошего здоровья и обладают множеством функций. Витамины A, D, E и K — жирорастворимые витамины — в основном содержатся в пищевых продуктах, содержащих жиры. Некоторые жирорастворимые витамины (например, витамин А) также содержатся в естественно обезжиренных продуктах, таких как зеленые листовые овощи, морковь и брокколи. Эти витамины лучше всего усваиваются в сочетании с жирными продуктами. Жиры также увеличивают биодоступность соединений, известных как фитохимические вещества, которые являются компонентами растений, такими как ликопин (содержится в томатах) и бета-каротин (содержится в моркови).Считается, что фитохимические вещества способствуют укреплению здоровья и благополучия. В результате, употребление помидоров с оливковым маслом или заправкой для салатов облегчит всасывание ликопина. Другие важные питательные вещества, такие как незаменимые жирные кислоты, являются составными частями самих жиров и служат строительными блоками клетки.
Рисунок 4.2.3: Пищевые источники жирорастворимых витаминов
Обратите внимание, что удаление липидных элементов из пищи также снижает содержание в ней жирорастворимых витаминов.При переработке таких продуктов, как зерно и молочные продукты, эти важные питательные вещества теряются. Производители заменяют эти питательные вещества с помощью процесса, называемого обогащением.
Инструменты для перемен
Помните, что жирорастворимым питательным веществам для эффективного усвоения необходим жир. Для следующего перекуса поищите продукты, содержащие витамины A, D, E и K. Есть ли в этих продуктах жиры, которые помогут вам их усвоить? Если нет, подумайте о том, как добавить немного полезных жиров, чтобы улучшить их усвоение.
Роль липидов в пище: источник высокой энергии
Продукты, богатые жирами, от природы имеют высокую калорийность.Продукты с высоким содержанием жира содержат больше калорий, чем продукты с высоким содержанием белка или углеводов. В результате продукты с высоким содержанием жиров являются удобным источником энергии. Например, 1 грамм жира или масла обеспечивает 9 килокалорий энергии по сравнению с 4 килокалориями в 1 грамме углеводов или белков. В зависимости от уровня физической активности и потребностей в питании потребности в жирах сильно различаются от человека к человеку. Когда потребность в энергии высока, организм приветствует высокую калорийность жиров.Например, младенцы и растущие дети нуждаются в достаточном количестве жира для поддержания нормального роста и развития. Если младенцу или ребенку давать диету с низким содержанием жиров в течение длительного периода, рост и развитие не будут нормально развиваться. Другие люди с высокими энергетическими потребностями — это спортсмены, люди, выполняющие тяжелую физическую работу, и те, кто выздоравливает после болезни.
Когда организм использует все свои калории из углеводов (это может произойти всего после двадцати минут упражнений), он начинает потребление жира.Профессиональный пловец должен потреблять большое количество пищевой энергии, чтобы соответствовать требованиям плавания на длинные дистанции, поэтому есть богатая жирами пища имеет смысл. Напротив, если человек, ведущий малоподвижный образ жизни, ест такую же жирную пищу, он будет потреблять больше жировых калорий, чем требуется их организму, всего за несколько укусов. Соблюдайте осторожность — потребление калорий сверх энергетической потребности является фактором ожирения.
Роль липидов в пище: запах и вкус
Жир содержит растворенные соединения, которые придают аппетитный аромат и вкус и улучшают вкусовые качества пищи.Жир также придает еде текстуру. Выпечка получается мягкой и влажной. При жарке продукты сохраняют вкус и сокращают время приготовления. Сколько времени вам нужно, чтобы вспомнить запах вашего любимого блюда? Какой была бы еда без этого пикантного аромата, который доставил бы вам удовольствие и повысил вашу готовность к еде?
Жир играет еще одну важную роль в питании. Жир способствует насыщению или ощущению сытости. Когда жирная пища проглатывается, организм реагирует, позволяя процессам, контролирующим пищеварение, замедлять движение пищи по пищеварительному тракту, тем самым способствуя общему ощущению сытости.Часто, прежде чем наступает чувство сытости, люди злоупотребляют жирной пищей, находя восхитительный вкус непреодолимым. Действительно, именно то, что делает жирную пищу привлекательной, также делает ее препятствием для поддержания здорового питания.
Инструменты для перемен
Хотя жиры придают нашим продуктам восхитительный запах, вкус и текстуру, они также содержат большое количество калорий. Чтобы позволить вашему телу ощутить эффект сытости от жира, прежде чем вы переедете, попробуйте смаковать жирную пищу.Медленное питание позволит вам полностью насладиться ощущением и насытиться меньшей порцией. Не забывайте не торопиться. Пейте воду между укусами или ешьте нежирную пищу до и после более жирной. Продукты с низким содержанием жира обеспечат объем, но с меньшим количеством калорий.
Ключевые выводы
Липиды включают триацилглицерины, фосфолипиды и стерины.
Триацилглицерины, наиболее распространенный липид, составляют большую часть жировых отложений и описываются в пищевых продуктах как жиры и масла.
Избыточная энергия пищи хранится в теле в виде жировой ткани.
Жиры имеют решающее значение для поддержания температуры тела, смягчения жизненно важных органов, регулирования гормонов, передачи нервных импульсов и сохранения памяти.
Липиды переносят жирорастворимые питательные вещества и фитохимические вещества и способствуют биодоступности этих соединений.
Жир — удобный источник энергии для людей с высокими энергетическими потребностями.


 С жирами в организм
С жирами в организм
 Известно, что нарушения метаболизма липидов возникают, например, как
Известно, что нарушения метаболизма липидов возникают, например, как
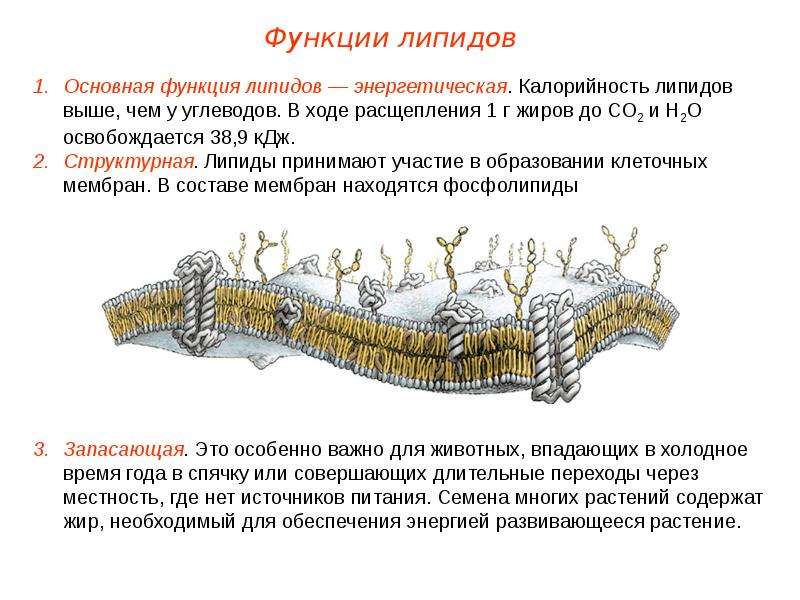 Протоплазматический (конститутивный) жир входит в состав всех структур клеток органов и тканей и в течение всей жизни остается практически на одном уровне – около 25 % общего жира организма.
Протоплазматический (конститутивный) жир входит в состав всех структур клеток органов и тканей и в течение всей жизни остается практически на одном уровне – около 25 % общего жира организма. Достижения химии липидов позволяют создавать пищевые добавки на основе природных липидов, улучшающих консистенцию и повышающих биологическую ценность продуктов.
Достижения химии липидов позволяют создавать пищевые добавки на основе природных липидов, улучшающих консистенцию и повышающих биологическую ценность продуктов. При полном сгорании 1 г липидов выделяется более чем в два раза больше энергии по сравнению с белками и углеводами. Запасные (резервные) липиды играют роль энергетического депо и локализуются в виде специализированных тканей (подкожная клетчатка, сальник, околопочечная капсула и т. п.). Они принимают участие в процессах терморегуляции, предохраняют кожу от высыхания, защищают органы от сотрясения, обеспечивают всасывание из кишечника жирорастворимых витаминов, являются потенциальным резервом эндогенной воды в организме (при окислении 100 г жира образуется 100 см3 воды) и др.
При полном сгорании 1 г липидов выделяется более чем в два раза больше энергии по сравнению с белками и углеводами. Запасные (резервные) липиды играют роль энергетического депо и локализуются в виде специализированных тканей (подкожная клетчатка, сальник, околопочечная капсула и т. п.). Они принимают участие в процессах терморегуляции, предохраняют кожу от высыхания, защищают органы от сотрясения, обеспечивают всасывание из кишечника жирорастворимых витаминов, являются потенциальным резервом эндогенной воды в организме (при окислении 100 г жира образуется 100 см3 воды) и др. В целом к липидам относят природные органические соединения, нерастворимые в воде, но растворимые в органических растворителях (бензол, бензин, петролейный эфир, этиловый эфир, ацетон, хлороформ, сероуглерод, метиловый и этиловый эфиры и т. п.). Пищевые липиды легко утилизируются животными организмами.
В целом к липидам относят природные органические соединения, нерастворимые в воде, но растворимые в органических растворителях (бензол, бензин, петролейный эфир, этиловый эфир, ацетон, хлороформ, сероуглерод, метиловый и этиловый эфиры и т. п.). Пищевые липиды легко утилизируются животными организмами.
 К ним также относят глико-, сульфолипиды и некоторые другие.
К ним также относят глико-, сульфолипиды и некоторые другие.
 Учитывая важное физиологическое значение и эссенциальность отдельных представителей липидов в обеспечении здорового питания, весьма важны их качественный состав и химическая природа.
Учитывая важное физиологическое значение и эссенциальность отдельных представителей липидов в обеспечении здорового питания, весьма важны их качественный состав и химическая природа. к. рНоптимум ее действия находится в слабощелочной среде. Однако природные тонкодиспергированные (эмульгированные) жиры (молока, яичного желтка) в желудке расщепляются.
к. рНоптимум ее действия находится в слабощелочной среде. Однако природные тонкодиспергированные (эмульгированные) жиры (молока, яичного желтка) в желудке расщепляются.
 к. глицерин водорастворим и свободно всасывается и поступает в кровь. Жирные кислоты переходят в растворимое состояние путем предварительного образования холиновых комплексов, представляющих собой соединения 2–4 парных желчных кислот и одной жирной кислоты. Все желчные кислоты (стероидные монокарбоновые оксикислоты) образуются в печени из холестерина. Они (главным образом холевая и дезоксихолевая кислоты) способны взаимодействовать с гликоколом (глицином) или таурином с образованием пептидоподобной связи. Такие соединения называют парными желчными кислотами.
к. глицерин водорастворим и свободно всасывается и поступает в кровь. Жирные кислоты переходят в растворимое состояние путем предварительного образования холиновых комплексов, представляющих собой соединения 2–4 парных желчных кислот и одной жирной кислоты. Все желчные кислоты (стероидные монокарбоновые оксикислоты) образуются в печени из холестерина. Они (главным образом холевая и дезоксихолевая кислоты) способны взаимодействовать с гликоколом (глицином) или таурином с образованием пептидоподобной связи. Такие соединения называют парными желчными кислотами.
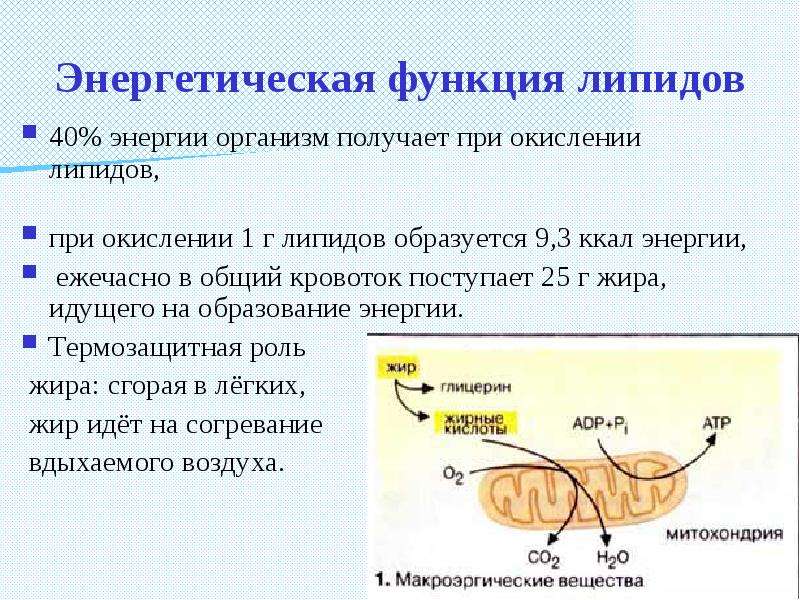 Они переносятся к различным органам и тканям, где подвергаются специфическим превращениям.
Они переносятся к различным органам и тканям, где подвергаются специфическим превращениям.



 Нарушения жирового обмена отмечаются и при недостатке в организме липотропных веществ типа холина, метионина, витамина F (фактора F), которые предохраняют печень от повышенного отложения жира путем участия в синтезе фосфолипидов.
Нарушения жирового обмена отмечаются и при недостатке в организме липотропных веществ типа холина, метионина, витамина F (фактора F), которые предохраняют печень от повышенного отложения жира путем участия в синтезе фосфолипидов.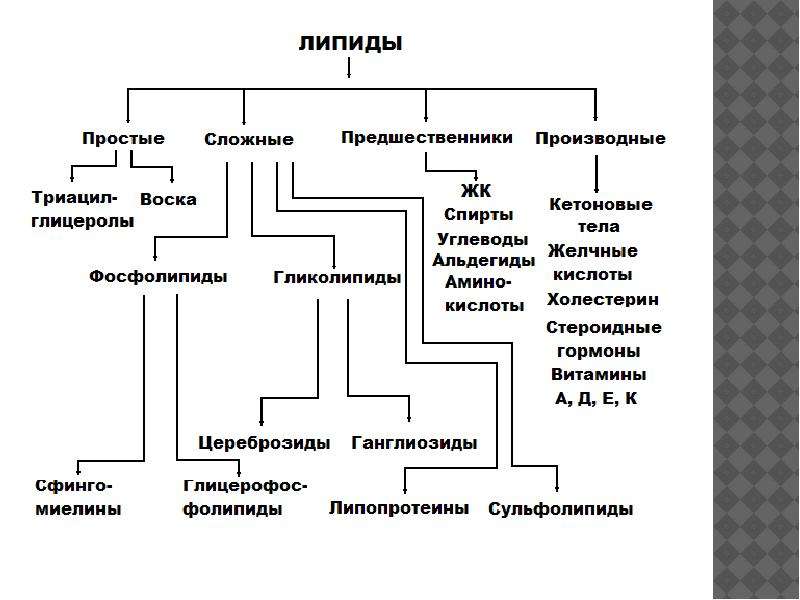 При избытке холестерина развиваются различные атеросклерозы.
При избытке холестерина развиваются различные атеросклерозы. К тому же полиненасыщенные жирные кислоты (ПНЖК), не синтезируемые в организме человека, относят к незаменимым (линолевая, линоленовая, арахидоновая). Арахидоновая кислота образуется в организме из линолевой при участии витамина В6. Линолевая образует и другие ПНЖК. По расположению первой двойной связи относительно метильной группы в молекуле ПНЖК относят к группам омега-6 (линолевая, γ-линоленовая, арахидоновая) и омега-3 (α-линоленовая, эйкозапентаеновая, докозагексаеновая).
К тому же полиненасыщенные жирные кислоты (ПНЖК), не синтезируемые в организме человека, относят к незаменимым (линолевая, линоленовая, арахидоновая). Арахидоновая кислота образуется в организме из линолевой при участии витамина В6. Линолевая образует и другие ПНЖК. По расположению первой двойной связи относительно метильной группы в молекуле ПНЖК относят к группам омега-6 (линолевая, γ-линоленовая, арахидоновая) и омега-3 (α-линоленовая, эйкозапентаеновая, докозагексаеновая).
 Суточная потребность здорового взрослого человека в фосфатидах составляет 5–10 г.
Суточная потребность здорового взрослого человека в фосфатидах составляет 5–10 г. к. они в равной степени необходимы человеку. Животные жиры – единственный источник витаминов А и D, растительные – витаминов Е и β-каротина.
к. они в равной степени необходимы человеку. Животные жиры – единственный источник витаминов А и D, растительные – витаминов Е и β-каротина. Они исключительно богаты жирорастворимыми витаминами и провитаминами – токоферолами, стероидами и каротиноидами, в их составе много водорастворимых витаминов – тиамина, рибофлавина, пиридоксина, биотина, фолиевой кислоты, пантотеновой кислоты, инозита, аскорбиновой кислоты.
Они исключительно богаты жирорастворимыми витаминами и провитаминами – токоферолами, стероидами и каротиноидами, в их составе много водорастворимых витаминов – тиамина, рибофлавина, пиридоксина, биотина, фолиевой кислоты, пантотеновой кислоты, инозита, аскорбиновой кислоты.
 Соотношение их в растительных тканях 3:1.
Соотношение их в растительных тканях 3:1. С его помощью получают вкусную, калорийную, с привлекательным внешним видом продукцию, которая пользуется большим спросом у потребителя.
С его помощью получают вкусную, калорийную, с привлекательным внешним видом продукцию, которая пользуется большим спросом у потребителя. 2).
2).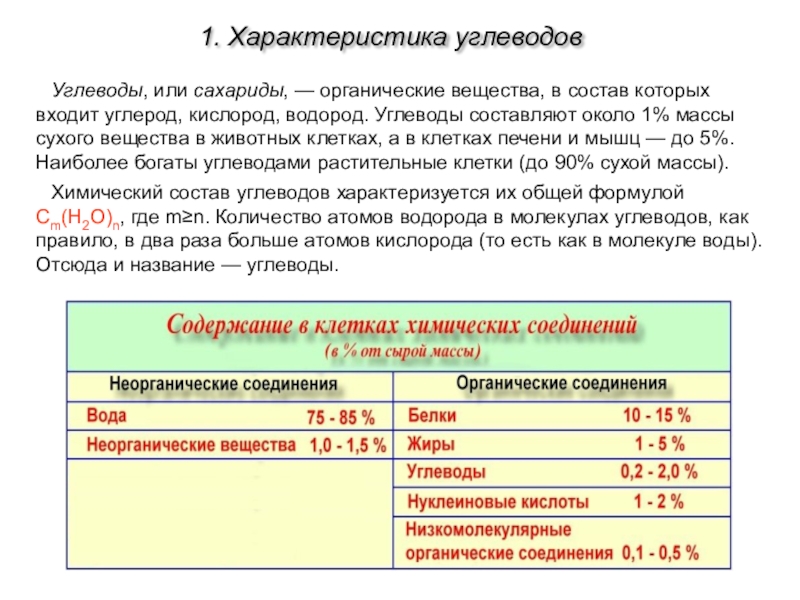

 Пищевые кислоты накапливаются
Пищевые кислоты накапливаются Применяются также соли
Применяются также соли Соли яблочной
Соли яблочной В
В Среднем считается норма в 102 г в сутки. В питании имеет значение не только количество, но и химический состав жиров. Линолевая и линоленовые кислоты не синтезируются в организме человека, арахидоновая кислота синтезируется из линолевой кислоты при участии витамина В6. Поэтому они получили название незаменимых или эсенциальных жирных кислот. В последние годы часто употребляется термин «полиненасыщенные жирные кислоты семейства омега – 3», в эту группу входят ά – линоленовая, эйкозапентаеновая, докозагексаеновая кислоты, содержащие несколько кратных связей и «полиненасыщенные жирные кислоты семейства омега – 6», в эту группу вхадит арахидоновая кислота.
Среднем считается норма в 102 г в сутки. В питании имеет значение не только количество, но и химический состав жиров. Линолевая и линоленовые кислоты не синтезируются в организме человека, арахидоновая кислота синтезируется из линолевой кислоты при участии витамина В6. Поэтому они получили название незаменимых или эсенциальных жирных кислот. В последние годы часто употребляется термин «полиненасыщенные жирные кислоты семейства омега – 3», в эту группу входят ά – линоленовая, эйкозапентаеновая, докозагексаеновая кислоты, содержащие несколько кратных связей и «полиненасыщенные жирные кислоты семейства омега – 6», в эту группу вхадит арахидоновая кислота. Повышенной биологической активностью обладают арахидоновая и линолевые кислоты. Среди продуктов питания наиболее богаты полиненасыщенными жирными кислотами растительные масла. Арахидоновая кислота содержится в яйцах, субпродуктах. Сбалансированный состав ежедневного рациона человека должен содержать 10 – 20 % полиненасыщенных жирных кислот, 50 – 60 % мононенасыщенных жирных кислот, 30 % насыщенных жирных кислот. \это обеспечивается при использовании в рационе одной трети растительных и двух третей животных жиров.
Повышенной биологической активностью обладают арахидоновая и линолевые кислоты. Среди продуктов питания наиболее богаты полиненасыщенными жирными кислотами растительные масла. Арахидоновая кислота содержится в яйцах, субпродуктах. Сбалансированный состав ежедневного рациона человека должен содержать 10 – 20 % полиненасыщенных жирных кислот, 50 – 60 % мононенасыщенных жирных кислот, 30 % насыщенных жирных кислот. \это обеспечивается при использовании в рационе одной трети растительных и двух третей животных жиров.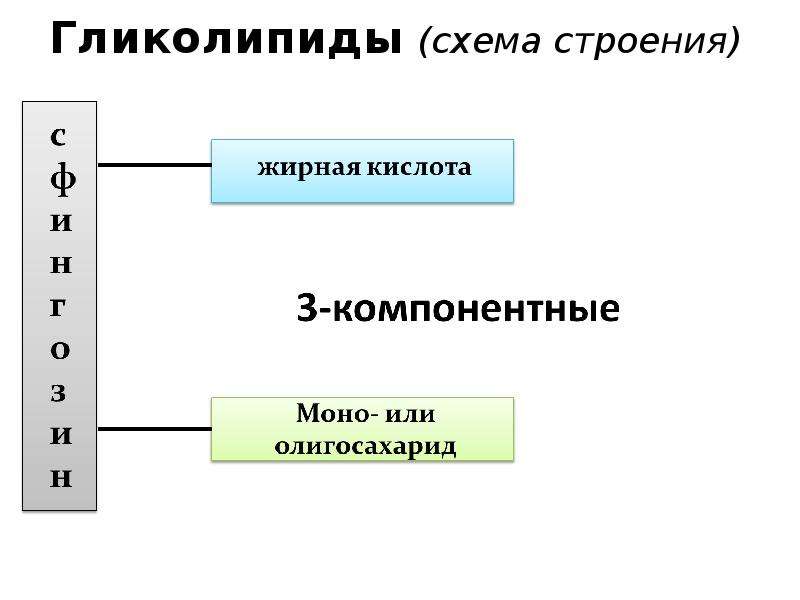
 Особую группу составляют жиры морских млекопитающих и рыб.
Особую группу составляют жиры морских млекопитающих и рыб.