

Что важно знать о витаминно-минеральных комплексах
Статті
18/12/2015
Даже сбалансированный рацион может быть на 20–30% дефицитным по некоторым витаминам [1]. При этом организм современного человека испытывает дефицит не только отдельных витаминов, но и минеральных веществ, а также их сочетаний. Причинами витаминно-минерального дефицита могут быть нерациональное питание, инфекционные и эндокринные заболевания, прием некоторых лекарств, неблагоприятное экологическое воздействие, интенсивные физические и эмоциональные нагрузки, период беременности и др. [2]. В то же время при проведении коррекции витаминно-минерального дефицита возникает ряд вопросов, требующих ответа
Возможно ли взаимодействуют ли компоненты в витаминно-минеральных комплексах?
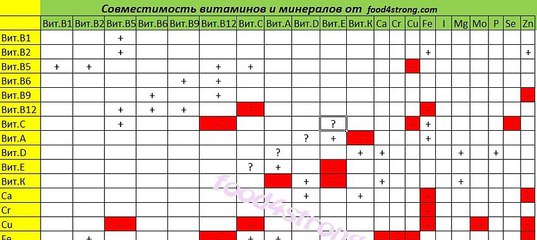
Известно около двух десятков взаимодействий витаминов и минералов — как положительных (синергизм), так и отрицательных (антагонизм).
В качестве примеров синергизма можно привести сочетание витамина D3 и кальция, витамина С и железа, витамина А и цинка, витамина В6 и магния.
С увеличением количества компонентов в витаминно-минеральном комплексе (ВМК) возрастают возможность и число взаимодействий между ингредиентами, что приводит к тому, что для многих компонентов начинают реализовываться отрицательные эффекты их действия друг на друга на разных уровнях [3]. В частности, витамин С окисляет до 30% витамина В12, который плохо совместим с витамином А, а также железом и медью. Фолиевая кислота образует нерастворимое соединение с оксидом цинка, что уменьшает усвоение витамина В9. При этом установлено, что многие минеральные вещества на этапах всасывания в кишечнике конкурируют друг с другом: кальций — за всасывание с железом, медью, магнием и свинцом; магний — за всасывание с железом, цинком и свинцом; медь — за всасывание с цинком, кальцием и кадмием; фосфаты ухудшают всасывание кальция, магния, меди и свинца; железо конкурирует за всасывание с кальцием, магнием, свинцом, фосфатами, цинком и кадмием, кроме того, железо является антагонистом цинка [1, 3].
Читайте также: Витамин D: вновь в центре внимания
Следует отметить, что применение монокомпонентных препаратов далеко не всегда является целесообразным, так как изолированный гиповитаминоз встречается достаточно редко — как правило, преобладает полигиповитаминоз. Таким образом, максимально эффективным витаминно-минеральным комплексом следует считать препарат, объединяющий синергические эффекты при исключении антагонизма микронутриентов [1].
Как решается проблема витаминно-минеральной несовместимости?
В процессе разработки и производства витаминно-минеральных комплексов учитывают варианты взаимодействия витаминов и минералов. К примеру, синергизм побуждает производителя объединить ингредиенты в одной таблетке, а антагонизм — вынуждает идти на изготовление отдельных гранул витаминов и минеральных веществ, а затем объединять их в обычную таблетку, двухслойную или ламинированную. Наиболее реакционноспособные компоненты могут быть включены в состав покрытия таблетки или в капсульную оболочку. Второй вариант решения проблемы состоит в «разведении» конкурирующих компонентов по разным таблеткам.
Второй вариант решения проблемы состоит в «разведении» конкурирующих компонентов по разным таблеткам.
Читайте также: Лекарственные взаимодействия: когда количество не переходит в качество
Следует отметить, что в твердых лекарственных формах легче избежать взаимодействия, используя некоторые витамины (например, цианокобаламин), заключенные в желатин, вместо чистой субстанции. Для инъекционных лекарственных форм витаминно-минеральных комплексов существует несколько методов предотвращения нежелательного взаимодействия между витаминами и минералами: использование двухкамерных ампул и лиофилизация (вымораживание). Для препаратов, предназначенных для перорального применения, целесообразным является приготовление порошков или растворимых гранул. Снижение содержания воды также способствует уменьшению вероятности нежелательных взаимодействий [2].
Совместимы ли витаминно-минеральные комплексы с другими лекарственными средствами?
При назначении витаминов на фоне применения других лекарственных средств необходимо учитывать возможности и особенности их взаимодействия.
К примеру, применение витаминов на фоне приема некоторых антибиотиков может уменьшать количество побочных явлений, усиливать их бактериостатическое действие и улучшать переносимость. В то же время применение витаминов группы В (особенно В1) повышает шокогенность антибиотиков пенициллинового ряда, а также левомицетина, амидопирина и новокаина. Не рекомендовано применять витаминно-минеральные комплексы одновременно с тетрациклинами. Витамин С, ввиду своего высокого окислительно-восстановительного потенциала, может изменять химический состав других препаратов. Аскорбиновая кислота повышает концентрацию в крови бензилпенициллина и тетрациклинов, а также снижает эффективность гепарина, непрямых антикоагулянтов и антибиотиков. При одновременном применении с ацетилсалициловой кислотой повышается выведение с мочой аскорбиновой кислоты и уменьшается экскреция ацетилсалициловой кислоты. Прием аскорбиновой кислоты повышает риск развития кристаллурии при лечении салицилатами и сульфаниламидами короткого действия, а также усиливает выведение препаратов, имеющих щелочную реакцию. Витамин В6 в терапевтических дозах снижает эффективность леводопы. Также это касается взаимодействия с циклосерином, адреналином, норадреналином и сульфаниламидами [2].
Витамин В6 в терапевтических дозах снижает эффективность леводопы. Также это касается взаимодействия с циклосерином, адреналином, норадреналином и сульфаниламидами [2].
Таким образом, курс витаминно-минеральной профилактики предпочтительно начинать после основного курса приема химиотерапевтических средств и, в частности, антибиотиков. Если в процессе применения витаминно-минеральных комплексов показан прием тетрациклинов, то интервал между приемом этих двух средств должен составлять не менее 2 ч. Во избежание передозировки не рекомендован одновременный прием витаминно-минеральных комплексов с другими препаратами, содержащими витамины и минералы [2].
Каковы особенности приема витаминно-минеральных комплексов в период беременности?
Недостаток витаминов и минералов в период беременности может отрицательно сказываться на здоровье не только самой женщины, но и плода, повышая риск перинатальной патологии и соответственно детской смертности, а также частоты недоношенности, врожденных уродств, нарушений физического и умственного развития детей.
Рост плода и увеличение собственной массы тела материнского организма, необходимого для обеспечения развития плода, требует дополнительных затрат энергии, в связи с чем потребность организма беременной в витаминах и минералах резко возрастает. Следует отметить, что восполнение недостатка витаминов путем дополнительного несбалансированного приема продуктов зачастую приводит к неадекватному увеличению потребления пищевых веществ и энергии, что в свою очередь нередко влечет за собой избыточное увеличение массы тела как беременной, так и плода. При этом зачастую усвоение витаминов из лекарственных средств выше, чем из пищевых продуктов, в которых они, как правило, находятся в связанной форме [1].
Читайте также: Кому и зачем нужна фолиевая кислота?
Особенно повышена потребность организма беременных в таких микронутриентах, как железо, фолиевая кислота, кальций, цинк и витамины С, В2, В6, В12 [1].
Не следует забывать, что прием специализированных витаминно-минеральных комплексов в ряде случаев предотвращает и/или уменьшает выраженность гестационного токсикоза. В частности, для профилактики явлений токсикоза, а также для коррекции угрожающего прерывания беременности в ранние сроки используют препараты, содержащие магний и витамин B6 [4].
В частности, для профилактики явлений токсикоза, а также для коррекции угрожающего прерывания беременности в ранние сроки используют препараты, содержащие магний и витамин B6 [4].
Читайте также: Болезни адаптации: токсикоз
Основным требованием к ВМК для беременных является прежде всего безопасность при максимальной усвояемости компонентов препарата [1].
Какие витаминно-минеральные комплексы назначают детям?
Широкое назначение витаминно-минеральных комплексов в педиатрии начинается при достижении ребенком возраста одного года. Проведение курса витаминно-минеральной коррекции необходимо в периоды повышенного расхода витаминов и минералов; по окончании курса антибиотикотерапии; в кризисные периоды развития иммунной системы [2].
Для детей младшего возраста важны сбалансированный состав и оптимальные дозы ингредиентов; минимизация аллергических реакций; удобство приема для ребенка и уверенности родителей в том, что ребенок принял препарат.
С учетом возможных взаимодействий витаминов и минералов составе витаминно-минеральных комплексов не должен быть перегружен по принципу «чем больше — тем лучше». Напротив, он должен быть оптимально сбалансирован, особенно в отношении минералов — детям до 2 лет рекомендованы поливитаминные препараты без минеральных веществ, т.к. последние (особенно микроэлементы) приводят к активации ферментов, что нецелесообразно в столь раннем возрасте [2]. С трехлетнего возраста возможно увеличение количества витаминов и включение в состав витаминно-минеральных комплексов йода, магния, цинка, селена и др.
Читайте также: Дети и лекарства
В свою очередь, периоды интенсивных нагрузок на детский организм требуют подбора определенного витаминно-минерального комплекса. В таких случаях для быстрой компенсации имеющегося дефицита эссенциальных микронутриентов необходимы дозы выше среднесуточных [4].
Современный педиатр должен иметь в арсенале линейку средств, характеризующуюся разнообразием лекарственных форм и возможностью назначения определенного препарата в профилактической или лечебной дозе. Кроме того, препарат должен обладать приятными органолептическими свойствами, что обусловливает комплаентность его приема [5].
Кроме того, препарат должен обладать приятными органолептическими свойствами, что обусловливает комплаентность его приема [5].
Подготовила Александра Демецкая,
канд. биол. наук
Список литературы находится в редакции
“Фармацевт Практик” #12′ 2015
Поділіться цим з друзями!
%PDF-1.6
%
1 0 obj
>
endobj
4 0 obj
/ModDate (D:20160506125259+03’00’)
/Subject
>>
endobj
2 0 obj
>
stream
application/pdf
Библиотека УО «ВГМУ»2016-05-06T12:52:59+03:002016-05-06T12:52:59+03:002016-05-06T12:52:59+03:00uuid:14f2e550-a97e-496b-81a9-a341e7e21f4fuuid:55b7ae49-b9a6-4d13-9f17-19febe15afc5
endstream
endobj
3 0 obj
>
endobj
5 0 obj
>
>>
/Rotate 0
/Type /Page
/Annots [20 0 R]
>>
endobj
6 0 obj
>
/Rotate 0
/Type /Page
>>
endobj
7 0 obj
>
/Rotate 0
/Type /Page
>>
endobj
8 0 obj
>
/Rotate 0
/Type /Page
>>
endobj
9 0 obj
>
/Rotate 0
/Type /Page
>>
endobj
10 0 obj
>
/Rotate 0
/Type /Page
>>
endobj
11 0 obj
>
/Rotate 0
/Type /Page
>>
endobj
12 0 obj
>
/Rotate 0
/Type /Page
>>
endobj
13 0 obj
>
/Rotate 0
/Type /Page
>>
endobj
14 0 obj
>
stream
xZ[sGv~跥`v:3~TRIe7|
| Эффекты при одновременном применении с препаратами | |
| Ванкомицин (бактерицидные антибиотики) | Синергизм. |
| Сульфаниламиды (бактериостатические антибиотики) | Антагонизм (амоксициллин действует только на размножающиеся микроорганизмы). |
| Аминогликозиды (бактерицидные антибиотики) | Синергизм. |
| Макролиды (бактериостатические антибиотики) | Антагонизм (амоксициллин действует только на размножающиеся микроорганизмы). |
| Фенилбутазон | Подавление канальцевой секреции препаратов пенициллинового ряда, что приводит к увеличению T1/2 и концентрации амоксициллина в плазме крови. |
| Пероральные контрацептивы | Возможно уменьшение эффективности пероральных контрацептивов. |
| Тетрациклины (бактериостатические антибиотики) | Антагонизм (амоксициллин действует только на размножающиеся микроорганизмы).
|
| Сульфинпиразон | Подавление канальцевой секреции препаратов пенициллинового ряда, что приводит к увеличению T1/2 и концентрации амоксициллина в плазме крови. |
| Ацетилсалициловая кислота | Подавление канальцевой секреции препаратов пенициллинового ряда, что приводит к увеличению T1/2 и концентрации амоксициллина в плазме крови. |
| Пробенецид | Уменьшение канальцевой секреции амоксициллина, что может сопровождаться повышением его концентрации в плазме крови. |
| Хлорамфеникол (бактериостатические антибиотики) | Антагонизм (амоксициллин действует только на размножающиеся микроорганизмы). |
| Циклосерин (бактерицидные антибиотики) | Синергизм. |
| Аллопуринол | Возможно появление кожной сыпи (механизм этого явления мало изучен). |
| Метотрексат | Возможно уменьшение клиренса метотрексата, по-видимому, вследствие уменьшения его канальцевой секреции в присутствии пенициллинов.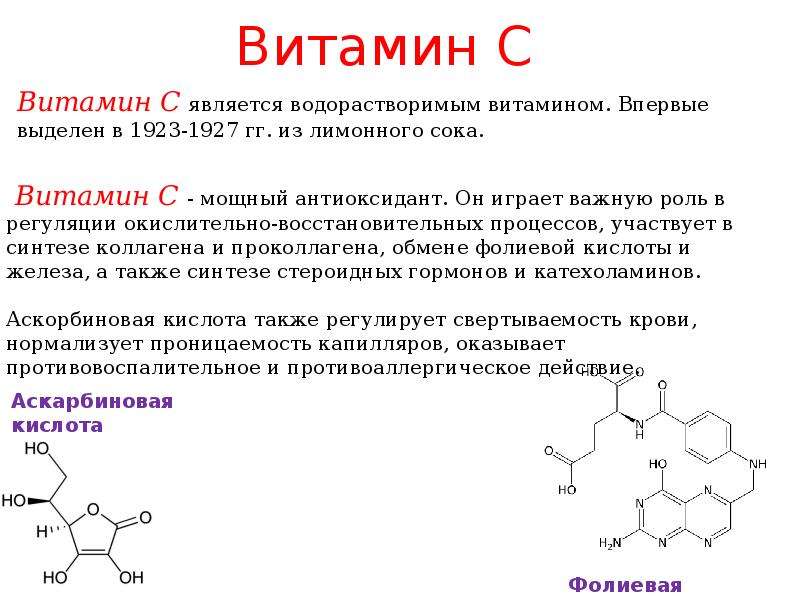 Описаны случаи усиления токсического действия метотрексата. Описаны случаи усиления токсического действия метотрексата.
|
| Цефалоспорины (бактерицидные антибиотики) | Синергизм. |
| Рифампицин (бактерицидные антибиотики) | Синергизм. |
| Аскорбиновая кислота | Повышение абсорбции амоксициллина. |
| Ланзопразол | Возможны глоссит, стоматит и/или появление темной окраски языка. |
| Оксифенбутазон | Подавление канальцевой секреции препаратов пенициллинового ряда, что приводит к увеличению T1/2 и концентрации амоксициллина в плазме крови. |
| Линкозамиды (бактериостатические антибиотики) | Антагонизм (амоксициллин действует только на размножающиеся микроорганизмы). |
| Непрямые антикоагулянты | Вследствие подавления под влиянием антибиотика кишечной микрофлоры, при участии которой осуществляется синтеза витамина К, возможно усиление антикоагулянтного действия снижение протромбинового индекса.
|
 ru
ru
Синергизм в косметике
Синергизм. Это слово произошло от греческого syneros и означает содействие, сотрудничество. Если мы обратимся к различным толковым словарям, то узнаем, что:
• в медицине термином синергизм обозначают совместное функционирование различных органов;
• в фармакологии – совокупное действие лекарственных веществ, каждое их которых усиливает эффект действия другого;
• в генетике и биологии – взаимное действие разнообразных физиологических и биохимических факторов, которое обуславливает оптимальный конечный эффект;
• в политологии — комбинированное действие на политические, экономические, социальные организации, при котором итоговый эффект превосходит действие, которое совершается каждым компонентом в отдельности.
То есть синергизм – это совместное действие элементов системы в одном направлении, усиление одного элемента другим элементом.
Надо учитывать и то, что помимо синергизма между ингредиентами возможно возникновение и отрицательного взаимного влияния (антагонизма), которое может привести к ослаблению эффектов от каждого из компонентов.
Рассмотрим эту проблему на косметических кремах.
Все ингредиенты, входящие в состав косметических средств, можно отсортировать по нескольким группам:
1. Вещества, которые создают структуру крема — эмульсию. В первую очередь, это растительные масла, которые вводятся в воду, тщательно перемешиваются (гомогенизируются) до однородного состояния. Если частички масла равномерно распределены в воде, то тогда обеспечивается хорошая впитываемость крема в кожу, в том числе и масляных составляющих, которые при обыкновенном нанесении не очень-то стремятся проникнуть в кожу. В настоящий момент существует оборудование, на котором процесс гомогенизации достиг высочайшего уровня, и на котором можно производить нанокрема с размером частиц меньше 15,7 нм. За счет малого размера частицы преодолевают барьер проницаемость кожи, проникают глубже основной массы клеточных слоев кожи и достигают уровня сосудистой (капиллярной) сети. А именно в частичках эмульсии крема как в растворителе или переносчике и содержатся все другие ингредиенты.
За счет малого размера частицы преодолевают барьер проницаемость кожи, проникают глубже основной массы клеточных слоев кожи и достигают уровня сосудистой (капиллярной) сети. А именно в частичках эмульсии крема как в растворителе или переносчике и содержатся все другие ингредиенты.
Помимо растительных масел в состав эмульсии вводятся и другие компоненты: эмоленты, стабилизаторы, загустители, эмульгаторы,- которые вводятся в состав продукта по технологической необходимости, обеспечивают легкость процесса эмульгирования, стабильность эмульсии.
2. Эстетические добавки – красители, ароматизаторы.
3. Вещества, необходимые для «долгожительства» косметических продуктов – консерванты.
Все выше упомянутые вещества являются базой для косметических кремов. Те задачи и функции, которые возлагаются на крем, выполняют так называемые активные компоненты. В основном это витамины, растительные экстракты, растительные и эфирные масла, вытяжки, антиоксиданты. Чтобы крем был более эффективным, необходимо чтобы он содержал всё это разнообразие видов активных ингредиентов, по 2 – 5 наименований по каждому виду. Чем разнообразнее состав активных составляющих крема, тем питательнее он будет для клеток кожи. При выборе компонентов надо учитывать эффект синергизма, который позволит не только сделать косметическое средство более питательным, но и уменьшить процент их ввода в состав крема.
Чтобы крем был более эффективным, необходимо чтобы он содержал всё это разнообразие видов активных ингредиентов, по 2 – 5 наименований по каждому виду. Чем разнообразнее состав активных составляющих крема, тем питательнее он будет для клеток кожи. При выборе компонентов надо учитывать эффект синергизма, который позволит не только сделать косметическое средство более питательным, но и уменьшить процент их ввода в состав крема.
Синергизм витаминов.
Примером синергизма может служить взаимодействие витаминов. В состав косметических кремов обычно входит несколько витаминов. Например, витамины С и Е практически всегда входят в состав кремов парой, они очень хорошо усваиваются, дополняя и усиливая действия друг друга.
Их сподвижником является витамин А. Витамин Е оберегает витамин А от окисления в тканях, поэтому в состав косметической продукции они, как правило, вводятся парой. Комбинация витаминов А и Е дает исключительный защитный и оздоровительный эффект, отсрочивает старение и делает возможным создание мощной антиоксидантной защиты.
Витамин В5, который принимает участие в регуляции липидного обмена, особенно результативен в сочетании с витаминами С, В2, D.
Синергизм антиоксидантов
Антиоксиданты — это вещества, которые охраняют организм от свободных радикалов и активных форм кислорода. Это главные стражи молодости и здоровья.
Они вводятся в косметические крема для выполнения сразу нескольких задач:
• для предотвращения окисления масел, которые входят в состав кремов, содержат полиненасыщенные жирные кислоты и которые имеют тенденцию легко окисляться;
• для повышения выработки коллагена и, как следствие, улучшения эластичности кожи;
• для эффективного снижения повреждений от солнечных лучей (устраняют фактор фитостарения).
Самые популярные антиоксиданты: витамины А, С, Е, РР, К, β-каротин (провитамин А), флавоноиды, танины, антоцианы, биофлавоноиды, коэнзим Q10, серосодержащие аминокислоты, селен и другие, а также растительные и ягодные экстракты таких растений как алоэ, зеленый чай, конский каштан, черника, эдельвейс, розмарин, облепиха, красный виноград, гранат, клюква, сосна, рябина, гингко билоба, гамамелис и многих-многих других.
При взаимодействии молекулы антиоксиданта со свободными радикалами первая теряет электрон, разрушается и «выходит из игры». Поэтому нужны восстанавливающие средства, которые переведут «испорченную» молекулу антиоксиданта в рабочее состояние. Витамин С «возвращает жизнь» витамину Е, но сам при этом окисляется. Соединения, содержащие серу, «реанимируют» витамин С, а биофлавоноиды активно восстанавливают и витамин Е, и витамин С. Подобный синергизм существует и промежду витамином Е и каротиноидами, промежду витамином Е и селеном (витамин Е необходим для сохранения биологических форм селена в активном состоянии). Равным образом селен снижает потребность в витамине Е, поддерживая его уровень в крови.Коэнзим Q10 (убихинон), представляющий собой витаминоподобный фермент, обладает антиоксидантными свойствами, уменьшает разрушение коллагена, является генератором энергии, снижает утерю кожей гиалуроновой кислоты, а в синергизме с витамином Е работает еще эффективнее, сильнее.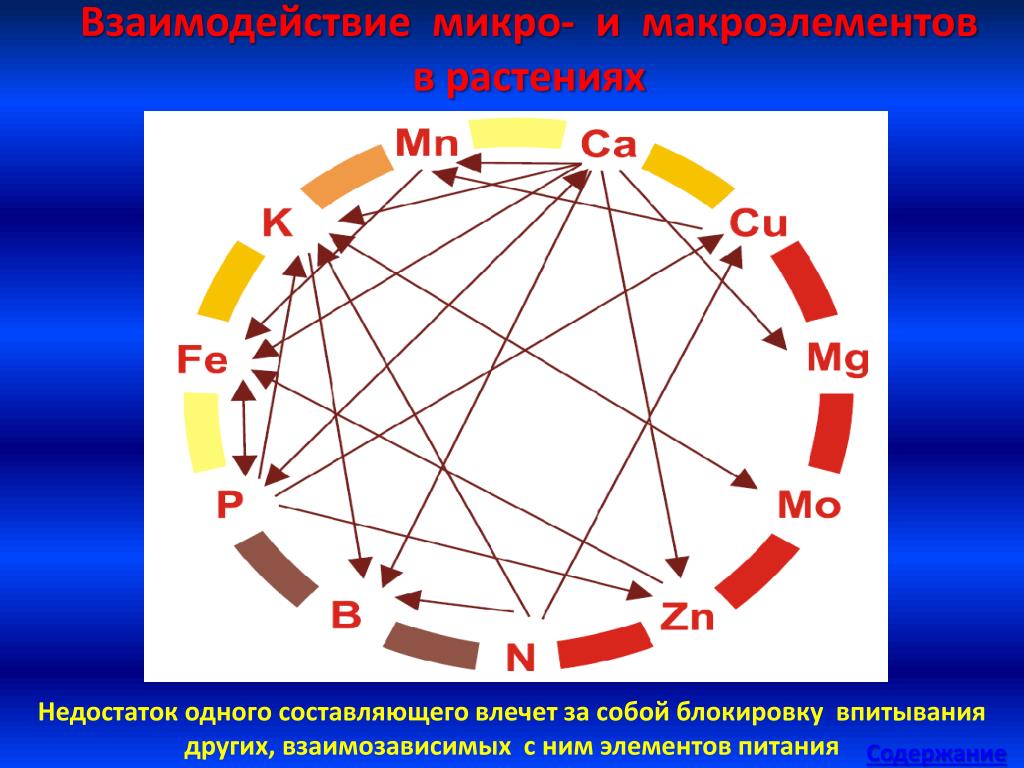 Таким образом синергизм антиоксидантов дает возможность достичь максимальной защиты и высокой стабильности продукта при более низкой концентрации антиоксидантов.
Таким образом синергизм антиоксидантов дает возможность достичь максимальной защиты и высокой стабильности продукта при более низкой концентрации антиоксидантов.
Большой сегмент среди активных ингредиентов косметических кремов занимают масла. Они помогают решить многие проблемы, возникающие с кожей: одни из них обеспечивают питанием сухую кожу и увлажняют ее, другие восстанавливают работу сальных желез, сужают поры, третьи – тонизируют кожу, делают ее упругой, эластичной. Продолжать можно и дальше.
Растительные масла – кладовая микроэлементов, витаминов, ненасыщенный жирных кислот, которые так необходимы для кожи.
Список масел, которые вводятся в косметические средства, огромен, все они обладают различными свойствами. И, конечно, хочется, чтобы смесь масел, которую мы вводим в крем, сочетала в себе сразу несколько свойств. Существуют так называемые базовые масла, процентное содержание которых должно быть в смеси больше. Они легко впитываются, не забивают поры. А есть дополнительные масла. И так же, как в случае с витаминами, антиоксидантами, необходимо учитывать синергизм масел, когда они помогают друг другу, и антагонизм, когда масла препятствуют положительному взаимному действию.
А есть дополнительные масла. И так же, как в случае с витаминами, антиоксидантами, необходимо учитывать синергизм масел, когда они помогают друг другу, и антагонизм, когда масла препятствуют положительному взаимному действию.
При покупке кремов и других косметических продуктов технологам всегда надо помнить как о синергизмевводимых компонентов, чтобы получить желаемый эффект при меньших материальных затратах, так и об антагонизме ингредиентов, чтобы не получить обратного результата: отрицательное взаимодействие вводимых компонентов и, как следствие, снижение эффективности косметического средства.
Диетотерапия пациентов с посттромботической болезнью на фоне приема варфарина
О центре —
Учимся быть здоровыми
18. 08.2020
08.2020
Автор: Manager
В настоящее время отмечается рост количества пациентов с посттромботической болезнью (ПТБ). Это одна из самых многочисленных групп, в нее входят пациенты с тромбозами, эмболиями и тромбофлебитами вен, из них 33 % принимают варфарин в течение длительного времени (от нескольких месяцев до нескольких лет).
В ежедневном рационе больного с посттромботической болезнью должно присутствовать большое количество сырых овощей и фруктов. В них много клетчатки, из которой в организме синтезируются фиброзные волокна, необходимые для «укрепления» венозной стенки. Следует ограничить прием жирных, острых и соленых блюд, которые за счет задержки жидкости могут привести к увеличению объема циркулирующей крови. Полезны продукты, содержащие жиры растительного происхождения.
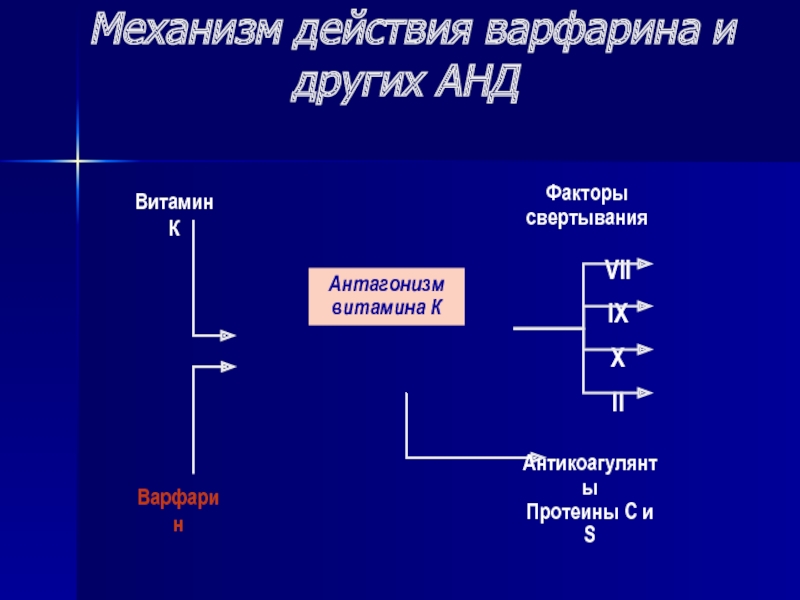
При приеме непрямых антикоагулянтов (варфарина) пациентами с ПТБ необходимо ограничивать в рационе продукты, которые содержат избыток витамина К. Их употребление входит в антагонизм с проводимой терапией. Варфарин снижает действие витамина К. Функция витамина К — обеспечение свертывания крови, что способствует остановке кровотечений из ран, и поддержание плотности костей человека.
Их употребление входит в антагонизм с проводимой терапией. Варфарин снижает действие витамина К. Функция витамина К — обеспечение свертывания крови, что способствует остановке кровотечений из ран, и поддержание плотности костей человека.
Недостаточность витамина К у человека приводит к замедлению свертывания и развитию геморрагического синдрома.
Алиментарный фактор не играет существенной роли в возникновении недостаточности витамина К вследствие широкого распространения витамина в пищевых продуктах и его термостабильности. Искусственно вызываемая недостаточность витамина К имеет место при применении антикоагулянтов непрямого действия.
Когда принимается варфарин, необходимо, чтобы витамин К содержался в рационе. Совсем исключать эти продукты тоже не рекомендуется. Недопустим прием алкоголя.
Однако важно, чтобы содержание витамина К в дневном рационе не менялось. Это будет способствовать улучшению действия варфарина. Если уровень витамина К в организме колеблется, то доктор будет вынужден часто менять дозировку препарата. Большинство людей при приеме варфарина могут питаться в своем обычном режиме. Но при этом важно помнить, что не следует употреблять значительно больше или значительно меньше, чем обычно ест пациент, продуктов, содержащих витамин К. Нужно следить за тем, чтобы уровень получаемого ежедневно витамина К был приблизительно одинаков.
Большинство людей при приеме варфарина могут питаться в своем обычном режиме. Но при этом важно помнить, что не следует употреблять значительно больше или значительно меньше, чем обычно ест пациент, продуктов, содержащих витамин К. Нужно следить за тем, чтобы уровень получаемого ежедневно витамина К был приблизительно одинаков.
Пища с высоким содержанием витамина К:
— Зеленые листовые овощи, например, капуста, шпинат, зеленая редька, швейцарский мангольд, листовая горчица, морская капуста и листья свеклы, горох;
— Брокколи, свежая или термически обработанная;
— Свежая петрушка;
— Зеленый чай;
— Авокадо, киви.
Пища со средним содержанием витамина К:
— Свежая листовая зелень, салат-латук, ромайн-латук, кочанный латук, а также цикорный салат, морские водоросли;
— Брюссельская капуста, цветная капуста, а также спаржа.
Витамин К также входит в состав многих поливитаминных комплексов, пищевых добавок и лекарственных сборов.
При приеме поливитаминов следует принимать их каждый день. Важно проконсультироваться с врачом по поводу приема пищевых добавок или лекарственных сборов. Некоторые из них могут содержать витамин К.
При употреблении с пищей витамина К намного больше, чем обычно, может снизиться эффект действия варфарина, что может привести к усилению тромбообразования. Получение с пищей витамина К намного меньше, чем обычно, может привести к усилению эффекта варфарина и повышению риска кровотечения.
Заведующая хирургическим отделением КДЦ № 1
Ольга Ивановна Свинтицкая
Питательные вещества | ASDhelp
ВЗАИМООТНОШЕНИЯ ПИТАТЕЛЬНЫХ ВЕЩЕСТВ
Минералы – витамины – эндокринные железы
Терапия о питании в значительной степени направлена на выявление и коррекцию дефицита питательных веществ. В настоящее время становится очевидным, что потеря гомеостатического равновесия между питательными веществами может оказать неблагоприятное воздействие на здоровье. Потеря этого жизненно важного баланса, особенно между микроэлементами, может привести к субклиническому дефициту.
Потеря этого жизненно важного баланса, особенно между микроэлементами, может привести к субклиническому дефициту.
Питательные взаимоотношения – сложные, особенно между микроэлементами. Один минерал не может быть поражен, без поражения, хотя бы, двух других минералов и т.д. Минеральные отношения можно сравнить с взаимосвязанными зубчатыми колесиками, некоторые связаны напрямую, некоторые косвенно. Любое движение одной шестеренки (минерала) скажется на движении всех других шестеренок (минералов). Степень и эффект каждой шестеренки (минерала) будет зависеть от его размера (количество минерала) и от количества зубцов в шестеренке (количество ферментов и биохимических реакций, в которых участвует минерал). Эта сеть шестерней выходит за пределы минеральных отношений, распространяясь и влияя на витамины, гормоны и неврологические функции.
Обширные исследования, которые включают в себя анализ минеральных тканей волос человека и других тканей, привели к значительному прогрессу в понимании отношений между минералами. Теперь эти знания могут применяться к отношениям между витаминами и эндокринным связям, что приводит к комплексному подходу к терапии о питании.
Теперь эти знания могут применяться к отношениям между витаминами и эндокринным связям, что приводит к комплексному подходу к терапии о питании.
Минеральные противоречия
Между микроэлементами существуют два вида взаимосвязей: антагонистическая и синергетическая, которые встречаются на двух уровнях – метаболический и абсорбционный. Сопротивление на абсорбционном уровне встречается из-за препятствующего поглощения; то есть избыточное потребление одного элемента может уменьшить кишечную абсорбцию другого элемента. Например, высокое потребление кальция снижает кишечную абсорбцию цинка, в то время как избыточное потребление цинка может снизить абсорбцию меди. Противоречия на метаболическом уровне появляются, если избыток одного элемента мешает метаболическим функциям другого или способствуют его выделению из-за частичной замены. Это наблюдается у цинка с медью, кадмия и цинка, железа и меди, кальция, магния и фосфора.
Минеральный синергизм
Взаимное усиление действия между элементами происходит, по большей части, на метаболическом уровне. Например, железо и медь синергетичны в этом, потому что для утилизации железа требуется медь. Магний также функционирует с калием, усиливая его задержку в клетках. Синергизм между кальцием, магнием и фосфором хорошо известен из-за их потребности в поддержке и структуре костной ткани. Другие минеральные синергизмы включают в себя:
Например, железо и медь синергетичны в этом, потому что для утилизации железа требуется медь. Магний также функционирует с калием, усиливая его задержку в клетках. Синергизм между кальцием, магнием и фосфором хорошо известен из-за их потребности в поддержке и структуре костной ткани. Другие минеральные синергизмы включают в себя:
Недостаточное потребление элементов может привести к накоплению токсичных соединений или других элементов.
При избыточном потреблении одного элемента, может возникнуть дефицит синергетического элемента.
Витаминные противоречия
Витамины также бывают с синергетическими и антагонистическими отношениями, которые рассматриваются не так часто. Противоречие могут быть непрямыми, они могут быть результатом чрезмерного употребления, могут повысить потребность в других витаминах. Примеры некоторых таких антагонизмов приведены ниже: Витамин А снижает токсичный эффект витамина D. Витамины А и D взаимно противоположны. Известно, что B1 может обладать антагонистичным действием на B12. Следует отметить, что антагонистические отношения между витамином С и В12 являются косвенными. Было подтверждено, что витамин С не действует напрямую на В12 и не разрушает его. Антагонизм связан с железом, так как известно, что железо противодействует кобальту, который является неотъемлемой частью витамина В12.
Известно, что B1 может обладать антагонистичным действием на B12. Следует отметить, что антагонистические отношения между витамином С и В12 являются косвенными. Было подтверждено, что витамин С не действует напрямую на В12 и не разрушает его. Антагонизм связан с железом, так как известно, что железо противодействует кобальту, который является неотъемлемой частью витамина В12.
Витамин С повышает усвоение железа, а значит косвенно влияет на состояние В12. Это, однако, редкое явление и может повлиять на небольшую часть населения, которая страдает от разных нарушений из-за перегрузки железом.
Витамин D усиливает всасывание кальция; поэтому, избыточное потребление витамина D за счет увеличения абсорбции кальция, приводит к пониженному сохранению в организме или к абсорбции магния, калия и фосфора. Эффект витамина А, который усиливает абсорбцию или удержание в организме калия и фосфора, что будет потом уменьшаться из-за высокого потребления витамина D.
Витаминный синергизм
Витамины вовлечены во множество реакций. Они действуют, как коферменты и синергически участвуют во многих ферментативных реакциях. Они также могут защищать организм от недостатка других витаминов. Ниже приведен список витаминных синергизмов:
Они действуют, как коферменты и синергически участвуют во многих ферментативных реакциях. Они также могут защищать организм от недостатка других витаминов. Ниже приведен список витаминных синергизмов:
Витаминно-минеральные синергизмы
Витамины тесно связаны с метаболическими функциями минералов. Хорошо известно, что дефицит витаминов может вмешиваться в утилизацию и абсорбцию минералов организмов, а добавки витаминов могут быть также нужны для устранения минерального дефицита. Классический пример потребности в витаминах и дефицита минералов являются рахиты и витамин D. Витамин С и\или В6 и витамин А могут потребоваться для коррекции анемии из-за дефицита железа, которая не будет реагировать на добавки железа. Дефицит цинка может быть связан с дефицитом витамина А, который не будет реагировать на добавку витамина А. Цинк нужен для мобилизации хранимого витамина А из печени. Ниже приведен список витаминно-минеральных синергизмов:
Витаминно-минеральные противоречия
Менее признаны антагонистические отношения между витаминами и минералами. Избыточное потребление определенного витамина может привести к нарушению минерала, посредством его дефицита или повышенной задержки этого минерала в организме. Высокое потребление витамина С может способствовать дефициту, как результату пониженной абсорбции или производства метаболических препятствий. Так как витамин С – антагонистичен к меди, а медь требуется в достаточном количестве для метаболической утилизации железа, повышенное потребление витамина С приведет к токсичности железа. Дефицит меди приводит к невозможности утилизации железа; поэтому железо будет накапливаться в запасающих тканях, если не будет достаточного количества меди. Медь и витамин С – синергетичны во многих метаболических функциях, из-за их антагонистического эффекта по отношению друг к другу, мы можем увидеть, что повышенное потребление меди может вызвать дефицит витамина С. Повышенное количество витамина С совместно с маргинальным статусом меди может содействовать образованию остеопороза так же, как и вызвать снижение иммунного ответа.
Избыточное потребление определенного витамина может привести к нарушению минерала, посредством его дефицита или повышенной задержки этого минерала в организме. Высокое потребление витамина С может способствовать дефициту, как результату пониженной абсорбции или производства метаболических препятствий. Так как витамин С – антагонистичен к меди, а медь требуется в достаточном количестве для метаболической утилизации железа, повышенное потребление витамина С приведет к токсичности железа. Дефицит меди приводит к невозможности утилизации железа; поэтому железо будет накапливаться в запасающих тканях, если не будет достаточного количества меди. Медь и витамин С – синергетичны во многих метаболических функциях, из-за их антагонистического эффекта по отношению друг к другу, мы можем увидеть, что повышенное потребление меди может вызвать дефицит витамина С. Повышенное количество витамина С совместно с маргинальным статусом меди может содействовать образованию остеопороза так же, как и вызвать снижение иммунного ответа. Повышенное потребление витамина D может привести к дефициту магния и калия, из-за их действия по увеличению абсорбции и\или удержанию кальция. Повышенное потребление витамина А может способствовать потере кальция.
Повышенное потребление витамина D может привести к дефициту магния и калия, из-за их действия по увеличению абсорбции и\или удержанию кальция. Повышенное потребление витамина А может способствовать потере кальция.
Питательно-эндокринные отношения
Эффекту, которое оказывает питательное действие на эндокринные железы, уделяется мало внимания. Известно, что гормоны влияют на питательные вещества на нескольких уровнях, включая абсорбцию, всасывание, перемещение и хранение. Питательные вещества, в свою очередь, могут оказать влияние на гормоны. Известно, что микроэлементы вовлечены в выделение гормонов, их активность и нацелены на связывающие ткани. Микроэлементы, в зависимости от их концентрации в организме (либо слишком большая, либо слишком маленькая) могут влиять на ось гипофиза гипоталамуса или на щитовидно-надпочечную ось.
Как с синергизмом и антагонизмом минералов и витаминов, эндокринный синергизм и антагонизм также существует.
Эндокринная классификация
Еще в 1940 году доктор Фрэнсис Поттенгер прокомментировал отношения между эндокринными железами и нервной системой. Позже доктор Мэлвин Пейдж блестяще классифицировал эндокринные железы в соответствии с неврологическим контролем, с симпатическим либо парасимпатическим. Он описал симпатическую группу, как «ускоренную» эндокринную, а парасимпатическую, как «замедленную». Симпатическая группа состоит из щитовидной железы, гормонов передней доли гипофиза, мозгового вещества надпочечников и андрогенных гормонов, произведенных яичниками. Парасимпатическая группа включает в себя поджелудочную железу, гормоны задней доли гипофиза, яичники, производящие эстрогены, паращитовидную железу и кору надпочечников. Доктор Пейдж выяснил, что если содержание фосфора в крови повышено, симпатическая группа является доминирующий, а если уровень кальция выше, чем уровень фосфора, то парасимпатическая группа является доминирующей. Он также заметил, что минеральное состав организма зависит непосредственно не от приема пищи, а от эффективности или неэффективности нейроэндокринной функции. Понимание этой работы д-ра Пейджа может помочь в классификации питательных веществ на две основные группы: симпатическая («ускоренная») или парасимпатическая («замедленная»).
Позже доктор Мэлвин Пейдж блестяще классифицировал эндокринные железы в соответствии с неврологическим контролем, с симпатическим либо парасимпатическим. Он описал симпатическую группу, как «ускоренную» эндокринную, а парасимпатическую, как «замедленную». Симпатическая группа состоит из щитовидной железы, гормонов передней доли гипофиза, мозгового вещества надпочечников и андрогенных гормонов, произведенных яичниками. Парасимпатическая группа включает в себя поджелудочную железу, гормоны задней доли гипофиза, яичники, производящие эстрогены, паращитовидную железу и кору надпочечников. Доктор Пейдж выяснил, что если содержание фосфора в крови повышено, симпатическая группа является доминирующий, а если уровень кальция выше, чем уровень фосфора, то парасимпатическая группа является доминирующей. Он также заметил, что минеральное состав организма зависит непосредственно не от приема пищи, а от эффективности или неэффективности нейроэндокринной функции. Понимание этой работы д-ра Пейджа может помочь в классификации питательных веществ на две основные группы: симпатическая («ускоренная») или парасимпатическая («замедленная»). Эти классификации основаны на питающие-эндокринных или эндокринно-питающие влиянии на нейроэндокринную функцию.
Эти классификации основаны на питающие-эндокринных или эндокринно-питающие влиянии на нейроэндокринную функцию.
Классификация питательных веществ с помощью эндокринного доминирования
Как говорит д-р Пейдж, фосфор может считаться симпатическим или стимулирующим. Кальций считается парасимпатическим или седативным. Симпатическая и парасимпатическая нейроэндокринные системы оказывают эффект на минералы, которые также могут классифицироваться, как стимулирующие или седативные, кроме кальция и фосфора.
Вместе с повышенным удержанием фосфора в организме, есть подобное повышение натрия и калия. С потерей кальция, обычно, происходит подобная потеря магния. Поэтому, фосфор, натрий и калий считаются симпатическими и стимулирующими питательными веществами.
Мы можем классифицировать некоторые основные минералы на симпатическую и парасимпатическую категория, из-за их нейроэндокринного влияния.
Витамины также могут классифицироваться в зависимости от их влияния на минеральный метаболизм и абсорбцию. Некоторые витамины и минералы. Как показано ниже, могут рассматриваться, как промежуточные, получается, что они могут производить, как стимулирующий эффект, так и седативный, в зависимости от их ферментативного и коферментного участия.
Некоторые витамины и минералы. Как показано ниже, могут рассматриваться, как промежуточные, получается, что они могут производить, как стимулирующий эффект, так и седативный, в зависимости от их ферментативного и коферментного участия.
Симпатическая и парасимпатическая классификация еды и воды
С пониманием нейроэндокринного влияния на питательные вещества, особенно микроэлементов, можно классифицировать любое вещество. Еда, вода, травы и лекарства – все попадают в стимулирующую (симпатическую) или седативную (парасимпатическую) категория. Еда и вода классифицируются в зависимости от их преобладающего минерального состава или ингибирующего абсорбционного эффекта. Классификация лекарств может основываться на их симпатомиметических, симпатолитических или парасимпатомиметических, парасимпатолитических эффектах, а также на их влиянии на метаболизм, абсорбцию и выделение.
Классификация еды
Вещества природного происхождения в еде могут препятствовать абсорбции минералов. Например, щавелевая кислота, которая найдена в таких продуктах, как шпинат, зелень свеклы и других продуктах, может сочетаться с кальцием в кишечном тракте, из-за чего она становится не абсорбируемой. Фитиновая кислота понижает усвоение кальция и абсорбцию цинка, она широко распространена в зерновых культурах и пшенице. Вымачивание этих продуктов понижает содержание кислоты и рекомендуется. Однако, смотря на их минеральное содержание, можно понять, что они богаты стимулирующими минералами, в отличие от седативных, и могут классифицироваться, как стимулирующие (симпатические) по своей природе. Минеральное содержание в еде варьируется в зависимости от почвы, на которой эта еда была выращена, а также от способов обработки еды и типа посуды, которая используема при приготовлении (медь, алюминий и т.д.).
Например, щавелевая кислота, которая найдена в таких продуктах, как шпинат, зелень свеклы и других продуктах, может сочетаться с кальцием в кишечном тракте, из-за чего она становится не абсорбируемой. Фитиновая кислота понижает усвоение кальция и абсорбцию цинка, она широко распространена в зерновых культурах и пшенице. Вымачивание этих продуктов понижает содержание кислоты и рекомендуется. Однако, смотря на их минеральное содержание, можно понять, что они богаты стимулирующими минералами, в отличие от седативных, и могут классифицироваться, как стимулирующие (симпатические) по своей природе. Минеральное содержание в еде варьируется в зависимости от почвы, на которой эта еда была выращена, а также от способов обработки еды и типа посуды, которая используема при приготовлении (медь, алюминий и т.д.).
Белковая еда
Белок обладает самым высоким специфическим динамическим действием (СДД), а поэтому вызывает повышение метаболической скорости (симпатомиметика). Часть эффектов происходит из-за кальция и магния, которые вырабатываются белком. Более плотные белки обладают большим СДД, чем белки с более низкой плотностью; говядина обладает более высоким СДД, чем рыба или птица, а овощные белки обладают самым низким СДД.
Более плотные белки обладают большим СДД, чем белки с более низкой плотностью; говядина обладает более высоким СДД, чем рыба или птица, а овощные белки обладают самым низким СДД.
Водяные растения
Жесткая вода, с высокой общей жесткостью, обычно, является щелочной. Седативные минералы, кальций и магний, обычно, в высоком содержании по отношению к стимулирующим минералам, и поэтому, считаются седативными (парасимпатическими).
Мягкая вода считается стимулирующей (симпатической), так как она обладает низким содержанием твердых веществ и, обычно, она является кислой, и при этом доминирующей в стимулирующих минералах, особенно в натрии. Использование трав всегда может быть более конкретным, в зависимости от их стимулирующего или седативного эффекта. Продолжающиеся исследования трав, показали, что в них содержится высокое содержание минеральных веществ, и классифицируются они соответствующим образом. Как пример седативного (парасимпатического) растения можно взять хвощ. его минеральное содержание кальция и магния – высоко, относительно натрия и калия. Как и с едой, минеральное содержание веществ в траве будет варьироваться в зависимости от почвы, на которой она была выращена.
его минеральное содержание кальция и магния – высоко, относительно натрия и калия. Как и с едой, минеральное содержание веществ в траве будет варьироваться в зависимости от почвы, на которой она была выращена.
Лекарства
Лекарства могут классифицироваться по их симпатомиметическому или парасимпатомиметическому действию, которые имитируют симпатическую или парасимпатическую активность нервной системы.
Некоторые симпатические лекарства включают в себя эпинефрин, фенилэфрин и метоксамин. Другие лекарства оказывают симпатическое действие, влияя на реализацию нейротрансмиттеров. К ним относятся эфедрин, тирамин и амфетамины. Эти лекарства обычно используются при лечении бронхиальных спазмов, связанных с проявлениями астмы и аллергии.
Симпатолитические препараты можно считать седативными, они центрально блокируют симпатическую активность, или поверхностно, ингибируя или блокируя нейротрансмиссию. Симпатические ингибиторы центрального действия включают в себя клонидин и метилдопа. Их часто встречающиеся названия на упаковках – Альдомет (Aldomet), Катапрес (Catapres), и Альдорил (Aldoril). Резерпин и раувольфия являются алкалоидами, которые препятствуют синтезу и хранению норэпинефрина, в то время как гуанетидин блокирует его реализацию. Некоторые торговые названия — Diupress, Harmonyl и Isme-lin. Блокираторы альфа- и бета- рецепторов – это празозин, феноксибензархин, пропанолол, надолол и метопролол. Их часто встречающиеся торговые названия Minipress, Dibenzyline, Lopressor, Corgard и Inderal. Эти препараты часто используются для лечения гипертонии.
Их часто встречающиеся названия на упаковках – Альдомет (Aldomet), Катапрес (Catapres), и Альдорил (Aldoril). Резерпин и раувольфия являются алкалоидами, которые препятствуют синтезу и хранению норэпинефрина, в то время как гуанетидин блокирует его реализацию. Некоторые торговые названия — Diupress, Harmonyl и Isme-lin. Блокираторы альфа- и бета- рецепторов – это празозин, феноксибензархин, пропанолол, надолол и метопролол. Их часто встречающиеся торговые названия Minipress, Dibenzyline, Lopressor, Corgard и Inderal. Эти препараты часто используются для лечения гипертонии.
Парасимпатические лекарства включают в себя: ацетилхолин, мускарин, пилокарпин, метахолин и карбамилхолин. Другие лекарства, которые усиливают эффект ацетохолина, являются неостигимин, физостигимин, пиридостигимин и хлорид карбамилметилихолина. Эти препараты часто используются для лечения неврологических и нейромускульных нарушений, как, например, миастения гравис. Для более полного списка симпатических и парасимпатических лекарств обратитесь к справочнику.
Лекарства, также препятствуют абсорбции и удержанию в организме питательных веществ. Например, антациды, слабительные средства, противосудорожные, кортикостероиды и антибактериальные средства, известны, как те, которые вызывают дефицит кальция и витамина D. Они оказывают хелатообразующее действие на кальций и противодействуют метаболическому эффекту витамина D. Длительное использование может привести к рахиту, остеомаляции и другим болезням, связанным с дефицитом кальция. Состояние питания человека может повлиять на метаболизм лекарств.
Классификация процессов болезней
Чтобы иметь возможность использовать вышесказанную информацию, мы должны знать о болезненных состояниях, которые проявляются, как симпатические или парасимпатические нарушения. Ниже будет приведен частичный список состояний, которые можно классифицировать соответствующим образом. Список составлен по результатам клинических исследований и по оценке более 100.000 профилей, которые были предоставлены врачами по всей стране. Этот список не должен рассматриваться, как полный или абсолютно точный, потому что всегда есть исключения. Для примера, гипертония может возникать и симпатически, и парасимпатически из-за различных факторов. Повышенная симпатическая стимуляция способствует гипертонии, но артерио- и атеро- склероз также может быть причиной гипертонии, либо симпатически, либо парасимпатически.
Этот список не должен рассматриваться, как полный или абсолютно точный, потому что всегда есть исключения. Для примера, гипертония может возникать и симпатически, и парасимпатически из-за различных факторов. Повышенная симпатическая стимуляция способствует гипертонии, но артерио- и атеро- склероз также может быть причиной гипертонии, либо симпатически, либо парасимпатически.
Пищевые индуцированные недостатки
Недостатки, полученные из-за питания (относительные или абсолютные), не редкие и часто появляются из-за повышенного употребления. Повышенное употребление, особенно отдельных питательных веществ, что иногда нужно организму, может вызвать фармакологическую реакцию. Реакцией на высокое потребление питательных веществ (витаминов и минералов) может быть вмешательство в использовании другого питательного вещества, что в итоге обладает противовитаминным или противоминеральным действием. Результат может быть благоприятным, но, если это будет продолжаться длительный период, может возникнуть дефицит другого питательного вещества. Например, чрезмерное потребление витамина Е может вызвать признаки и симптомы похожие на дефицит витамина А. добавка витамина А может противодействовать эффекту витамина Е и, в итоге, это приведет к дефициту витамина D. Эти побочные эффект можно предотвратить, уменьшив потребление витамина Е. Или другой пример, если пациент испытывает симптомы дефицита кальция и его организм не отвечает на 800 или 1000 мг-ые добавки кальция в день, первым делом в клиниках предложат увеличить дозы, в два или даже в три раза больше, чем сейчас. Это может только усугубить симптомы пациента, но даже после нескольких месяцев, сокращение в потреблении кальция может привести к незамедлительному возвращению всех симптомов. Чтобы сохранять у пациента бессимптомное состояние, дозы со временем будут увеличиваться, а не уменьшаться. Если рассматривать синергистов и антагонистов кальция, как добавки витамина D, магния или меди, и уменьшение потребления витамина Е, витамина А, калия, фитиновой и щавелевой кислоты, то организм пациента может реагировать только на 400 мг кальция в день.
Например, чрезмерное потребление витамина Е может вызвать признаки и симптомы похожие на дефицит витамина А. добавка витамина А может противодействовать эффекту витамина Е и, в итоге, это приведет к дефициту витамина D. Эти побочные эффект можно предотвратить, уменьшив потребление витамина Е. Или другой пример, если пациент испытывает симптомы дефицита кальция и его организм не отвечает на 800 или 1000 мг-ые добавки кальция в день, первым делом в клиниках предложат увеличить дозы, в два или даже в три раза больше, чем сейчас. Это может только усугубить симптомы пациента, но даже после нескольких месяцев, сокращение в потреблении кальция может привести к незамедлительному возвращению всех симптомов. Чтобы сохранять у пациента бессимптомное состояние, дозы со временем будут увеличиваться, а не уменьшаться. Если рассматривать синергистов и антагонистов кальция, как добавки витамина D, магния или меди, и уменьшение потребления витамина Е, витамина А, калия, фитиновой и щавелевой кислоты, то организм пациента может реагировать только на 400 мг кальция в день.
Заключение
Понимание питания и его важной роли в нашем здоровье постоянно развивается и становится более распространенным среди людей, как неотъемлемая часть медицинской помощи, особенно среди современных прогрессивных представителей медицинских услуг. В книге «Иммунитет и Инфекции через питание, Механизмы воздействия» Р. К. Чандра говорится: «…было обнаружено, что функция многих типов клеток изменяется при дефиците питательных веществ». Чандра говорит в своих наблюдениях, что не только недоедание, но и переедание могут изменить ответную реакцию иммунитета. Это особенно заметно благодаря микроэлементам, потому что слишком большое количество микроэлементов может быть таким же вредным, как и слишком малое. Абсолютный минеральный дефицит в наше время редкость, однако относительные дефицитные состояния встречаются часто. С лучшим пониманием и применением этой темы, можно осуществить более комплексный подход к своему здоровью, избегая «рулетку питательных веществ», которая была описана ранее. Конкретное применение известных стимулирующих и седативных веществ при индивидуальном лечении может улучшить реакцию организма при этом с меньшим количеством нежелательных побочных эффектов.
Конкретное применение известных стимулирующих и седативных веществ при индивидуальном лечении может улучшить реакцию организма при этом с меньшим количеством нежелательных побочных эффектов.
(PDF) Черных Н.А., Баева Ю.И. Антагонизм и синергизм
элементами могут носить антагонистический или
синергетический характер, что контролируется многими
факторами, механизмы действия которых еще плохо изучены.
В общем виде антагонизм и синергизм можно связать со
способностью одного элемента ингибировать или
стимулировать поглощение других элементов растениями.
При этом антагонистические эффекты реализуются двумя
путями: макрокомпонент ингибирует поглощение
микроэлемента, или, наоборот, микрокомпонент ингибирует
поглощение макрокомпонента. Однако следует подчеркнуть,
что все эти реакции весьма переменчивы — иногда и для
антагонистических пар наблюдаются синергические эффекты
[3]. К настоящему времени накоплен достаточно большой
К настоящему времени накоплен достаточно большой
экспериментальный материал по данной проблеме, однако
однозначных ответов на вопросы о характере
взаимодействия между различными химическими
элементами при поступлении в растения не получено.
Цель исследования — оценка изменения элементного
состава растений пшеницы и люцерны под воздействием
рахных уровней загрязнения почв тяжелыми металлами (Pb и
Сd).
Объекты исследования: пахотный горизонт (0-25 см)
дерново-подзолистой почвы (южная часть Московской обл.,
Ступинский район), стебли и листья яровой пшеницы сорта
«Московская 35» и люцерны «Вега-87».
Химические свойства почвы: рНводн. 5,6; содержание
гумуса 2,2%, сумма поглощенных оснований 7,1 мгэкв/100г;
емкость катионного обмена 14,9 мгэкв/100г.
Исследуемые виды растений: Пшеница (Triticum) — род
травянистых, в основном однолетних, растений семейства
Злаки, или Мятликовые (Poaceae), ведущая зерновая
культура во многих странах. Люцерна посевная, (Medicago
Люцерна посевная, (Medicago
sativa) — травянистое растение, типовой вид рода Люцерна
Минеральные и витаминные антагонисты и синергисты
Динамический баланс питательных веществ
В анализе минералов волосяных тканей интерпретируется уникальный минеральный профиль человека и разрабатывается план сбалансированного питания в контексте ДИНАМИЧЕСКИХ синергетических и антагонистических отношений, которые имеют минералы. к минералам и минералам приходится на витамин.
Эти взаимосвязи сложны, но их важно помнить, потому что, когда они игнорируются, дополнительные недостатки или дисбалансы могут быть созданы из-за добавок.Никогда не стоит смотреть только на абсолютные уровни минерала, чтобы составить план балансировки минералов. Напротив, само искусство интерпретации HTMA и балансировки минералов человека основано на рассмотрении и понимании этой динамики.
Дэвид Л. Уоттс, основатель лаборатории Trace Elements (лаборатории, которую я использую для всех волос моего клиента HTMA) объясняет это следующим образом:
«Минеральные отношения можно сравнить с серией взаимосвязанных механизмов, которые все связаны между собой, некоторые прямо, а некоторые косвенно.
Любое движение одной шестерни (минерала) приведет к перемещению всех других шестерен (минералов). Степень воздействия на каждую шестерню (минерал) будет зависеть от размера шестерни (количества минералов) и количества зубцов шестерни (количества ферментов или биохимических реакций, в которых участвует минерал). Эта взаимосвязь механизмов выходит за рамки только минеральных отношений, она распространяется на витамины, гормоны и неврологические функции и влияет на них ». (Источник)
Эти минеральные динамики устанавливались на протяжении многих лет не только в физиологии человека, но также в ветеринарии и здоровье почвы.Хотя влияние минералов на людей, животных и растения разное, каждое в зависимости от своего уникального метаболизма, динамика минералов по отношению друг к другу согласуется.
Волосы — это ткань выделения , которая сильно отличается от крови, которая представляет собой ткань , транспортирующую . Поэтому результаты лабораторных анализов волос не следует интерпретировать как результаты анализа крови.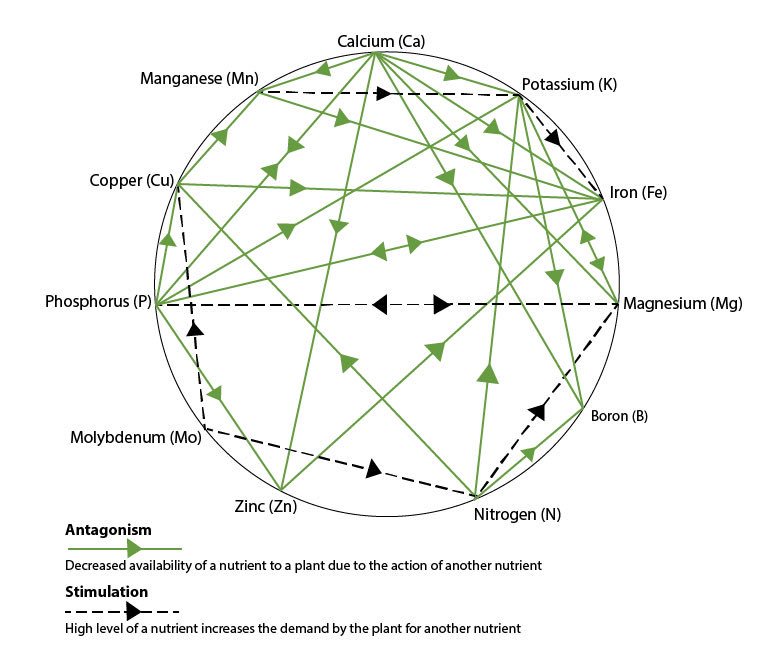 Результаты анализа крови и волос часто не совпадают не только потому, что они представляют собой совершенно разные временные рамки (1 час по сравнению с 3 месяцами), но и потому, что две ткани служат разным целям в организме.
Результаты анализа крови и волос часто не совпадают не только потому, что они представляют собой совершенно разные временные рамки (1 час по сравнению с 3 месяцами), но и потому, что две ткани служат разным целям в организме.
Тем не менее, минералы действительно обнаруживаются в волосах в соответствии с последовательными и последовательными узорами, тест за тестом. Значения очень точные, а не произвольные. При хорошем понимании динамики минералов из лаборатории HTMA можно многое узнать о метаболизме человека, помимо абсолютных цифр.
Многие слухи о неточности HTMA происходят из-за того, что врачи, заказывающие лаборатории HTMA, не всегда обучены интерпретировать лабораторные результаты с точки зрения динамики минералов.
Итак, сегодня я хочу поделиться некоторыми основными антагонистическими и синергетическими отношениями между минералами и витаминами. Клиенты просили меня предоставить эту информацию, и я надеюсь, что она послужит простой справкой и отправной точкой для понимания этих отношений.
Если вам интересно узнать больше о некоторых из наиболее распространенных минеральных дисбалансов и закономерностей при мигрени, прочтите эту статью.
Минеральный синергизм и антагонизм проявляются на двух уровнях: уровне абсорбции и уровне метаболизма внутри клетки.Например, избыток одного минерала может повлиять на всасывание другого в кишечном тракте, ИЛИ он может повлиять на клеточный метаболизм путем истощения другого минерала в клетке или и того, и другого.
В HTMA мы часто видим «дамп», возникающий при потере соты. Свалка — это недостаток на клеточном уровне, который отличается от недостатка, вызванного неспособностью усвоить минерал.
Есть много преимуществ, которые можно получить в исцелении, если мы рассмотрим баланс питательных веществ по отношению друг к другу и то, как они влияют на усвоение и метаболизм.Зная эти факторы, мы можем принимать антагонисты минералов в разное время дня для лучшего усвоения, и мы можем быть уверены, что не будем принимать слишком много одного витамина или минерала за счет другого.
Некоторые минералы являются одновременно синергическими и антагонистическими
Чтобы усложнить ситуацию, многие минералы действуют совместно в оптимальных соотношениях, но когда имеется избыток одного минерала, он может противодействовать минералу, с которым обычно работает согласованно. Цинк и медь — классический тому пример.Хотя они конкурируют за абсорбцию, и хотя избыток меди или цинка истощает другие, они также работают вместе во многих ферментативных процессах. Например, и цинк, и медь необходимы для выработки эстрогенов и антиоксидантного фермента SOD (супероксиддисмутазы).
Минеральные антагонисты
В HTMA мы используем минеральные круги в качестве наглядного пособия для обозначения антагонизма минералов. Ниже минеральное колесо.
Здесь вы можете увидеть антагонистическое отношение минералов друг к другу.
Как видите, у некоторых минералов есть стрелки, идущие в одном направлении, что означает, что минерал, на который указывает стрелка, опускается из-за минерала, от которого указывает стрелка. Например, фосфор снижает содержание натрия, но не натрия. Другие пары минералов имеют две стрелки, показывающие, что оба минерала антагонистичны друг другу, то есть кальций снижает фосфор, а фосфор снижает уровень кальция.
Например, фосфор снижает содержание натрия, но не натрия. Другие пары минералов имеют две стрелки, показывающие, что оба минерала антагонистичны друг другу, то есть кальций снижает фосфор, а фосфор снижает уровень кальция.
Когда мы видим низкие уровни минерала в лаборатории HTMA, мы затем пытаемся определить, вызван ли дефицит человека избытком других антагонистических минералов в таблице, или дефицит вызван плохой ассимиляцией всех минералов. из-за низкого уровня кислоты в желудке или минерального истощения из-за надпочечниковой недостаточности.Другие минералы в таблице, наряду с физиологическими симптомами от тщательного приема, дают важные подсказки о том, где и почему происходит потеря. Эти подсказки можно правильно интерпретировать только через понимание динамики минералов.
Например, человек с низким уровнем большинства питательных элементов с большей вероятностью будет иметь проблемы с усвоением минералов. Человек с повышенным уровнем одного минерала, такого как марганец, с одновременно очень низким уровнем меди, с большей вероятностью будет иметь дефицит меди, вызванный токсичностью марганца (поскольку марганец является мощным антагонистом меди). В этом случае мы знаем, что марганец вряд ли будет выражать клеточную потерю (или выведение, или «сброс»), потому что других минералов, которым марганец противодействует (магний, кальций и железо), также мало. С другой стороны, содержание натрия в лаборатории этого клиента намного выше, чем у большинства больных мигренью, но согласуется с тем, как марганец повышает уровень натрия. Если бы этот марганец выражал клеточную потерю, вероятно, в лаборатории был бы сигнал, указывающий, что вызывает его потерю, а других минералов, которые антагонистичны марганцу, было бы не так мало.
В этом случае мы знаем, что марганец вряд ли будет выражать клеточную потерю (или выведение, или «сброс»), потому что других минералов, которым марганец противодействует (магний, кальций и железо), также мало. С другой стороны, содержание натрия в лаборатории этого клиента намного выше, чем у большинства больных мигренью, но согласуется с тем, как марганец повышает уровень натрия. Если бы этот марганец выражал клеточную потерю, вероятно, в лаборатории был бы сигнал, указывающий, что вызывает его потерю, а других минералов, которые антагонистичны марганцу, было бы не так мало.
Следующая информация о минералах предоставляется для справки. Эти колеса, описания и взаимосвязи были разработаны Дэвидом Л. Уоттсом из лаборатории микроэлементов и взяты из этого справочника: Взаимосвязи между питательными веществами.
Кальций
Здесь вы можете увидеть антагонистические отношения кальция к минералам и витаминам.
Более девяноста процентов кальция в организме хранится в костях и зубах, которые действуют как резервуары, из которых может выводиться кальций, необходимый для внескелетных функций. Кальций содержится практически в каждой клетке по всему телу и считается биологическим мессенджером, ответственным за передачу сигналов для целевых действий с клетками через определенные кальциевые каналы. Кальций регулируется в тканях и сыворотке за счет структур скелета. Он регулируется паращитовидными железами и почками и зависит от инсулина, надпочечников, а также мужских и женских гормонов. Дисбаланс кальция относительно его синергических и антагонистических питательных веществ может быть основным фактором остеопороза даже при адекватном потреблении кальция с пищей.
Кальций содержится практически в каждой клетке по всему телу и считается биологическим мессенджером, ответственным за передачу сигналов для целевых действий с клетками через определенные кальциевые каналы. Кальций регулируется в тканях и сыворотке за счет структур скелета. Он регулируется паращитовидными железами и почками и зависит от инсулина, надпочечников, а также мужских и женских гормонов. Дисбаланс кальция относительно его синергических и антагонистических питательных веществ может быть основным фактором остеопороза даже при адекватном потреблении кальция с пищей.
Магний
Здесь вы можете увидеть антагонистические отношения магния к минералам и витаминам.
Магний является четвертым по распространенности катионом в организме и ключевым элементом клеточных метаболических функций. Магний отвечает за активацию более 200 критических ферментов. Из-за обширной роли магния в биологических процессах признание его синергетической и антагонистической роли с другими питательными веществами чрезвычайно важно и может значительно повысить его терапевтическую эффективность при многих состояниях здоровья.
Хром
Здесь вы можете увидеть антагонистические отношения хрома к минералам и витаминам.
Хром — важный микроэлемент, важный для переработки углеводов и жиров и помогающий клеткам правильно реагировать на инсулин. Известно, что хром является составной частью фактора толерантности к глюкозе (GTF) и действует синергетически с инсулином, способствуя усвоению глюкозы клетками. Хром важен для структуры и метаболизма нуклеиновых кислот. С статусом хрома связан ряд физиологических и болезненных состояний.
Медь
Здесь вы можете увидеть антагонистические отношения меди к минералам и витаминам.
Медь входит в состав многих ферментов, включая цитохром-с-оксидазу, супероксиддисмутазу, церулоплазмин, дофамин-B-гидроксилазу, лизилоксидазу и моноаминоксидазу. Дисбаланс меди по отношению к другим питательным веществам может нарушить активность этих важных функций ферментов. Следует отметить, что избыток меди столь же серьезен, как и ее дефицит. Антагонистические питательные вещества, показанные в следующей таблице, могут помочь снизить чрезмерное содержание меди в тканях.
Цинк
Здесь вы можете увидеть антагонистические отношения цинка к минералам и витаминам.
Цинк — еще один важный элемент, необходимый для активности более ста ферментов. Цинк участвует в иммунной регуляции, противовирусной активности, росте и развитии, и, возможно, его наиболее важная роль — потребность в цинке в синтезе РНК. Таким образом, баланс цинка с другими питательными веществами в организме имеет решающее значение для нормального здоровья, но оценка этого баланса имеет решающее значение при лечебном питании.
Железо
Здесь вы можете увидеть антагонистические отношения железа к минералам и витаминам.
Железо участвует во многих метаболических процессах, особенно в ферментах, и в результате многие клинические проявления, включая анемию, могут развиться в результате дефицита железа. В то время как анемия, конечно, является наиболее распространенным заболеванием, связанным с дефицитом железа, другие состояния, такие как: сидеропения может способствовать нарушениям иммуномодуляции, эндокринным, физическим и даже эмоциональным расстройствам. Избыток или токсичность железа может быть обнаружен на противоположном конце спектра состояния железа, и с ним можно конкретно бороться, предоставляя питательные вещества, антагонистические по отношению к железу.
Избыток или токсичность железа может быть обнаружен на противоположном конце спектра состояния железа, и с ним можно конкретно бороться, предоставляя питательные вещества, антагонистические по отношению к железу.
Марганец
Здесь вы можете увидеть антагонистические отношения марганца к минералам и витаминам.
Марганец находится в основном в митохондриях клеток. Следовательно, на структуру и функцию митохондрий особенно влияет статус марганца. Марганец не только отвечает за активацию митохондриальной супероксиддисмутазы, но также активирует ферменты, связанные с метаболизмом жирных кислот и синтезом белка, которые очень важны для нормального функционирования клеток.
Селен
Здесь вы можете увидеть антагонистические отношения селена к минералам и витаминам.
Была обнаружена прямая биохимическая роль селена в его связи с активностью глутатионпероксидазы. Синергетические отношения селена с витамином Е показали, что он является ингибитором химических канцерогенов, ускоряя их детоксикацию. Селен защищает от хромосомных повреждений, стимулирует восстановление ДНК и регулирует скорость деления клеток. С тех пор было обнаружено, что селен играет роль в нормальной экспрессии щитовидной железы и способствует периферическому превращению Т4 в Т3 в печени и почках.
Селен защищает от хромосомных повреждений, стимулирует восстановление ДНК и регулирует скорость деления клеток. С тех пор было обнаружено, что селен играет роль в нормальной экспрессии щитовидной железы и способствует периферическому превращению Т4 в Т3 в печени и почках.
Синергисты
Минеральные / минеральные синергисты
Витаминно-минеральные синергисты
Витаминно-витаминные синергисты
Хотите узнать, каков ваш персональный профиль минералов?
Надеюсь, этот справочник по антагонистам и синергистам минералов и витаминов был для вас полезным!
Общее понимание динамики минералов и витаминов становится еще более ценным, если применять его к пониманию вашего индивидуального профиля минералов.Вот где приходит HTMA! Если вы хотите составить собственный отчет HTMA, он предлагается вместе с моей курсовой работой, чтобы дать вам прекрасную основу и отправную точку для начала лечения мигрени.
Антагонист витамина B-6 из льняного семени нарушает метаболизм аминокислот у самцов крыс с умеренным дефицитом витамина B-6 | Журнал питания
Аннотация
Предпосылки: Пиридоксаль-5′-фосфат (PLP) играет решающую роль в качестве кофактора в метаболизме аминокислот. Среди населения широко распространен умеренный дефицит витамина B-6, который может усугубляться из-за приема внутрь 1-амино D-пролина (1ADP), антагониста витамина B-6, обнаруженного в семенах льна.
Среди населения широко распространен умеренный дефицит витамина B-6, который может усугубляться из-за приема внутрь 1-амино D-пролина (1ADP), антагониста витамина B-6, обнаруженного в семенах льна.
Цель: Учитывая предварительные данные о влиянии синтетического 1АДФ на показатели метаболизма пиридоксина, настоящее исследование было разработано для изучения влияния 1АДФ, полученного из льняного семени, на метаболизм аминокислот у крыс с умеренным дефицитом витамина B-6.
Методы: самцов крыс-отъемышей ( n = 8 / лечение) потребляли полуочищенный рацион, содержащий либо 7 мг гидрохлорида пиридоксина / кг рациона [оптимальный витамин B-6 (OB)], либо 0.7 мг гидрохлорида пиридоксина / кг диеты [умеренный дефицит витамина B-6 (MB)], каждый с 0 или 10 мг антагониста витамина B-6 / кг диеты, либо в синтетической форме (1ADP), либо в виде экстракта льняного семени (FE) , в течение 5 нед. В конце эксперимента были проанализированы концентрации витамина B-6 и аминокислот в плазме, а также активность печеночных PLP-зависимых ферментов.
Результаты: По сравнению с контрольной группой MB, концентрации PLP в плазме были на 26% и 69% ниже, соответственно, у крыс MB + FE и MB + 1ADP ( P ≤ 0.001). В группе MB + FE концентрация цистатионина в плазме была на 100% выше, а концентрации α-аминомасляной кислоты и глутаминовой кислоты в плазме были на 59% и 30% ниже, соответственно, чем в контрольной группе MB. Как синтетический 1ADP, так и FE значительно ( P <0,001) ингибировали in vitro печеночную активность 2 PLP-зависимых ферментов, цистатионин-β-синтазы (до 44%) и цистатионин-γ-лиазы (до 60%), независимо от концентрации витамина B-6. Из-за воздействия антагониста витамина B-6 наблюдаемые нарушения биомаркеров плазмы и активности печеночных ферментов не были очевидны или имели меньшую степень у крыс, потребляющих достаточное количество витамина B-6.
Заключение: Текущие данные модели на крысах свидетельствуют о том, что антагонист витамина B-6, преобладающий в настоящее время в пищевых продуктах человека, может создавать проблемы для людей с умеренным статусом витамина B-6.
Введение
Витамин B-6 известен своей биологической ролью кофактора в бесчисленных метаболических реакциях из-за его способности связывать различные ферментативные реакции углерода и азота и его участия в биосинтезе биогенных аминов и одноуглеродных единиц (1, 2).Обычно пиридоксаль-5′-фосфат (PLP) 7 , активная форма витамина B-6, связывается с ε-аминогруппой активных остатков лизина, содержащихся в витамин B-6-зависимых ферментах, образуя таким образом основание Шиффа / внешний альдимин. Последний вид затем действует как общий центральный промежуточный продукт для всех реакций, катализируемых PLP, включая декарбоксилирование, рацемизацию, трансаминирование, β-элиминирование и альдольное расщепление, и, таким образом, служит для выполнения различных функций в метаболизме аминокислот, сахаров и липидов (2 , 3).В некоторых метаболических путях аминокислот, включая пути транс-сульфирования и реметилирования метаболизма метионина, витамин B-6 необходим в качестве кофактора (4–6). В пути транс-сульфирования PLP действует как кофермент для цистатионин-β-синтазы (CBS), который конденсирует гомоцистеин с серином с образованием цистатионина и цистатионин-γ-лиазы (CGL), которая превращает цистатионин в цистеин, высвобождая α-аминомасляную кислоту в виде побочный продукт. В пути реметилирования гомоцистеин снова превращается в метионин путем связывания нескольких ферментов, включая серингидроксиметилтрансферазу (SHMT).Этот PLP-зависимый фермент участвует в обратимой реакции по переносу группы -CH 2 — от серина к тетрагидрофолату с образованием 5,10-метилентетрагидрофолата, причем последний также образуется через систему расщепления глицина (4, 5, 7, 8).
В пути транс-сульфирования PLP действует как кофермент для цистатионин-β-синтазы (CBS), который конденсирует гомоцистеин с серином с образованием цистатионина и цистатионин-γ-лиазы (CGL), которая превращает цистатионин в цистеин, высвобождая α-аминомасляную кислоту в виде побочный продукт. В пути реметилирования гомоцистеин снова превращается в метионин путем связывания нескольких ферментов, включая серингидроксиметилтрансферазу (SHMT).Этот PLP-зависимый фермент участвует в обратимой реакции по переносу группы -CH 2 — от серина к тетрагидрофолату с образованием 5,10-метилентетрагидрофолата, причем последний также образуется через систему расщепления глицина (4, 5, 7, 8).
Статус витамина B-6 в организме в основном определяется его диетическим питанием и может нарушаться приемом антагониста витамина B-6 (9). Некоторые препараты (изониазид, циклосерин, пеницилламин и местранол), а также природные вещества (гиромитрин, канаванин, агаритин, тиофиллин, кофеин и линатин) действуют как антагонисты витамина B-6 в организме, уменьшая свободный PLP и / или ингибируя активность ферментов пиридоксалькиназы (PLK) и пиридоксинфосфатоксидазы (PNPOx) (9–11).Линатин, встречающийся в природе дипептид глутаминовой кислоты и 1-амино D-пролина (1ADP), является антагонистом витамина B-6, присутствующим в льняном семени. Химически это 1 — [(n-γ-L-глутамил) амино] -D-пролин. После гидролиза соляной кислотой линатин высвобождает 1ADP, активную форму этого антагониста витамина B-6 (12). Предыдущие исследования документально подтвердили, что общее содержание антагонистов витамина B-6, выраженное в эквивалентах 1-амино-D-пролина (1АДФЭ; равно 1/2 линатина + 1АДФ), составляло 177–437 мкг / г цельного льняного семени в зависимости от сорта. выборка (12, 13).1ADP связывается с PLP с образованием комплексов гидразона, и эти комплексы могут уменьшать пул свободного PLP in vivo или вызывать токсичность гидразона в организме (12, 14). Ранее мы показали, что токсичность синтетического 1АДФ была гораздо более очевидной у животных, у которых был умеренный дефицит витамина B-6, по сравнению с животными, у которых был избыток. Пероральный прием синтетического 1АДФ в концентрации 10 мг / кг диеты вызывал серьезные нарушения метаболизма гомоцистеина у крыс с умеренным дефицитом витамина B-6 (15).
Потребление льняного семени растет из-за его предполагаемой пользы для здоровья, включая снижение факторов риска, связанных с диабетом, раком и сердечно-сосудистыми заболеваниями (16–18). Одновременный прием антинутриентов, включая антагонист витамина B-6 1ADP, присутствующий в льняном семени, населением в целом, особенно тем, у кого умеренный дефицит витамина B-6, может вызывать беспокойство. Поскольку явный дефицит витамина B-6 встречается редко и преобладает умеренный дефицит (19, 20), настоящее исследование было разработано для оценки влияния антагониста витамина B-6 из льняного семени на метаболизм аминокислот на модели грызунов с умеренным содержанием витамина B. -6 дефицит.
Методы
Приготовление экстракта льняного семени.
Льняное семя было приобретено на коммерческом рынке и обезжирено с использованием аппарата Сокслета с гексаном в качестве растворителя. Обезжиренное льняное семя повторно измельчали до получения тонкого порошка. Антагонист витамина B-6 из молотого обезжиренного льняного семени был экстрагирован с использованием метода ультразвуковой обработки, описанного ранее, с небольшими модификациями, включая использование 40% изопропанола при 25 ° C с соотношением растворителя к твердому веществу 10: 1. на 30 мин (13).Экстракт концентрировали при пониженном давлении и сушили вымораживанием. Затем его тонко измельчали, и количество общего содержания антагониста витамина B-6 было определено количественно с использованием сверхвысокопроизводительной жидкостной хроматографии / масс-спектрометрии с синтетическим 1ADP (Santa Cruz Biotechnology) и собственным линатином в качестве стандартов ( 13). Экстракт льняного семени (FE), содержащий концентрированный антагонист витамина B-6, хранили при -20 ° C до использования.
Животные и диета.
Сорок восемь крыс-самцов Sprague-Dawley массой 112 ± 9 г были приобретены в Центре ухода за животными Университета Манитобы.Их по отдельности помещали в полипропиленовые клетки в комнате, в которой поддерживалась температура 20 ± 2 ° C с 12-часовым ритмом света / темноты при относительной влажности 50–70%. После акклиматизации в течение 1 недели крыс случайным образом разделили на 6 групп ( n = 8) и кормили полуочищенным рационом (AIN-93G, на основе безвитаминного казеина), содержащим гидрохлорид пиридоксина (PN⋅HCl) в дозе 7 мг / кг. [оптимальный витамин B-6 (OB)] или 0,7 мг / кг диеты [умеренный дефицит витамина B-6 (MB)], используя модель умеренного дефицита витамина B-6, как описано ранее (15).Каждая группа крыс также потребляла ab libitum 10 мг / кг антагониста витамина B-6 либо в синтетической форме (1ADP) (Santa Cruz Biotechnology), либо без FE, либо без него (контроль) в течение 5 недель ( Supplemental Table 1 ). Прием пищи контролировали ежедневно, а массу тела измеряли каждую неделю. В конце эксперимента образцы плазмы и тканей были собраны после 12 часов голодания для биохимических анализов.
Биохимические анализы плазмы.
Витамеры B-6 в плазме, включая PLP, пиридоксаль, пиридоксин и 4-пиридоксиновую кислоту (4-PA), были измерены как их семикарбазидные производные с использованием ВЭЖХ и детекции флуоресценции, как описано ранее (15).Общие тиолы в плазме, включая гомоцистеин и цистеин, были количественно определены с использованием 7-фторбензо-2-окса-1,3-диазол-4-сульфоната аммония методом ВЭЖХ с детектированием флуоресценции (21). Анализ профиля аминокислот в плазме выполняли с использованием имеющегося в продаже набора (EZ: faast kit, Phenomenex) с использованием пары Varian 450-GC с Varian 240-MS IT.
Активность печеночных PLP-зависимых ферментов.
Образцы печени (0,5 г) гомогенизировали с 5 мл ледяного 50 ммоль / л калий-фосфатного буфера (pH 6.8) и центрифугировали при 15000 × g и 4 ° C для сбора супернатанта (15). Активность печеночного фермента CBS определяли с использованием ранее описанного метода с некоторыми модификациями. Образец супернатанта инкубировали в реакционной смеси, содержащей радиоактивный материал, 25 ммоль / л L- [U- 14 C] серина (~ 78000 дпм / мкмоль) (PerkinElmer) и L-цистатионин (0,15 ммоль / L), DL-гомоцистеин (41,67 ммоль / л), S -аденозилметионин (0,32 ммоль / л), DL-пропаргилглицин (2.08 ммоль / л), Трис (125 ммоль / л) и ЭДТА (2,08 ммоль / л). После инкубации при 37 ° C подсчитывали радиоактивность новообразованного 14 C-цистатионина для определения активности фермента CBS (22). Печеночную активность CGL определяли с помощью ферментно-связанного анализа с использованием ранее описанного спектрофотометрического метода (23).
Статистический анализ.
Данные были разрезаны на основе концентрации PN⋅HCl в рационе (OB и MB), чтобы определить простые эффекты воздействия антагонистов независимо от воздействия витамина B-6 с пищей, статистическая процедура, использованная в предыдущей работе (15).Затем был проведен статистический анализ с использованием SPSS 16.0. Нормальность данных проверяли перед анализом с помощью теста Шапиро-Уилка, и, при необходимости, данные нормализовали преобразованием log 10 . Общая линейная модель использовалась для сравнения эффектов лечения в той же группе с витамином B-6 с использованием теста Бонферрони, установив значимость на уровне P <0,05. Для всех биохимических аналитов плазмы мы использовали ANCOVA, взяв потребление корма в качестве ковариаты для контроля, учета и выравнивания несоответствий, которые могли возникнуть из-за различий в потреблении корма.В противном случае мы использовали однофакторный дисперсионный анализ для сравнения эффектов лечения.
Результаты
Препарат ИП.
При количественной оценке с помощью жидкостной хроматографии / масс-спектрометрии сверхвысокого разрешения было обнаружено, что количество антагониста витамина B-6 в лиофилизированных FE составляет 305 мкг линатина и 17 мкг 1ADP на 50 мг образца. Согласно молярному расчету, упомянутому в нашем предыдущем исследовании (13), общее количество антагониста витамина B-6, выраженное как 1ADPE, присутствующее в образце 50 мг, составляло 169 мкг, и это значение использовалось при приготовлении FE рационы, содержащие 10 мг 1АДФЭ / кг рациона.
Рабочие характеристики.
У крыс, получавших OB-рационы, те, которые подвергались воздействию 1ADP или FE, не демонстрировали каких-либо значительных различий в потреблении корма, конечной массе тела, коэффициенте эффективности корма (FER) или общем приросте веса по сравнению с крысами, получавшими контрольный OB-рацион. (Рисунок 1 и Таблица 1). Однако у крыс, получавших диету с MB, у крыс из группы MB + 1ADP было более низкое потребление корма, FER и общий прирост массы тела, чем у крыс, получавших контрольную диету с MB ( P <0.001). В результате окончательная масса тела также была значительно снижена ( P <0,001). Тем не менее крысы в группе MB + FE не отличались от контрольных крыс по маркерам показателей роста. Кроме того, кормление 1ADP, независимо от источника, не изменяло относительный вес печени крыс, потребляющих рационы с оптимальными или умеренно недостаточными концентрациями PN⋅HCl (таблица 1). В течение исследования наблюдалась одна летальность в группе OB + 1ADP.Смерть наступила внезапно, и у крысы не наблюдалось анорексии или снижения веса; однако патологоанатомическое исследование не выявило причину смерти.
РИСУНОК 1
Влияние FE и 1ADP на рост крыс, получавших рационы, содержащие 7 мг или 0,7 мг PN⋅HCl / кг рациона в течение 5 недель. Значения — это средние значения ± SEM; n = 8. В группе витамина B-6 обозначенные без общей буквы различаются, P <0,05. FE, экстракт льняного семени; МБ, умеренный дефицит витамина B-6; OB, оптимальный витамин B-6; PN⋅HCl, гидрохлорид пиридоксина; 1АДФ, 1-амино D-пролин.
РИСУНОК 1
Влияние FE и 1ADP на рост крыс, получавших рационы, содержащие 7 мг или 0,7 мг PN⋅HCl / кг рациона в течение 5 недель. Значения — это средние значения ± SEM; n = 8. В группе витамина B-6 обозначенные без общей буквы различаются, P <0,05. FE, экстракт льняного семени; МБ, умеренный дефицит витамина B-6; OB, оптимальный витамин B-6; PN⋅HCl, гидрохлорид пиридоксина; 1АДФ, 1-амино D-пролин.
ТАБЛИЦА 1
Влияние FE и 1ADP на массу тела, потребление корма и коэффициент эффективности корма у крыс, получавших рационы, содержащие PN⋅HCl либо в дозе 7 мг / кг, либо в дозе 0.7 мг / кг диеты в течение 5 недель 1
| Группа витамина B-6 (PN⋅HCl) и лечение . |
Начальная масса тела, г . |
Конечная масса тела, г . |
Общая прибавка в весе, г . |
RLW . |
Потребление корма, г . |
FER . |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OB диета (7 мг / кг диеты) | ||||||||||||||
| OB | 112 ± 6 | 456 256 3 .49 ± 0,13 | 930 ± 15,3 | 0,37 ± 0,01 | ||||||||||
| OB + FE | 113 ± 3 | 456 ± 14 | 343 ± 16 | 3,49 ± 0,06 | 918 ± 243,7 | |||||||||
| OB + 1ADP | 114 ± 3 | 430 ± 12 | 316 ± 12 | 3,57 ± 0,07 | 869 ± 24,8 | 0,37 ± 0,01 | ||||||||
|
4 | 9030 .26 | 0,2 | 0,79 | 0,29 | 0,56 | |||||||||
| MB диета (0,7 мг / кг диеты) | 383 ± 8 a | 270 ± 10 a | 3,44 ± 0,08 | 780 ± 15,8 a | 0,35 ± 0,01 a | |||||||||
| MB + FE | 2 | 112 ± 7 a | 260 ± 7 a | 3.35 ± 0,09 | 770 ± 18,8 a | 0,34 ± 0,01 a | ||||||||
| MB + 1ADP | 114 ± 2 | 255 ± 8 b | 141 ± 8 b ± 0,14 | 514 ± 19,0 b | 0,28 ± 0,02 b | |||||||||
| P | 0,98 | <0,001 | <0,001 | 0,72 0,001 | 001 |
| Группа витамина B-6 (PN⋅HCl) и лечение . |
Начальная масса тела, г . |
Конечная масса тела, г . |
Общая прибавка в весе, г . |
RLW . |
Потребление корма, г . |
FER . |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OB диета (7 мг / кг диеты) | ||||||||||||||
| OB | 112 ± 6 | 456 256 3 .49 ± 0,13 | 930 ± 15,3 | 0,37 ± 0,01 | ||||||||||
| OB + FE | 113 ± 3 | 456 ± 14 | 343 ± 16 | 3,49 ± 0,06 | 918 ± 243,7 | |||||||||
| OB + 1ADP | 114 ± 3 | 430 ± 12 | 316 ± 12 | 3,57 ± 0,07 | 869 ± 24,8 | 0,37 ± 0,01 | ||||||||
|
4 | 9030 .26 | 0,2 | 0,79 | 0,29 | 0,56 | |||||||||
| MB диета (0,7 мг / кг диеты) | 383 ± 8 a | 270 ± 10 a | 3,44 ± 0,08 | 780 ± 15,8 a | 0,35 ± 0,01 a | |||||||||
| MB + FE | 2 | 112 ± 7 a | 260 ± 7 a | 3.35 ± 0,09 | 770 ± 18,8 a | 0,34 ± 0,01 a | ||||||||
| MB + 1ADP | 114 ± 2 | 255 ± 8 b | 141 ± 8 b ± 0,14 | 514 ± 19,0 b | 0,28 ± 0,02 b | |||||||||
| P | 0,98 | <0,001 | <0,001 | 0,72 0,001 | 001 |
ТАБЛИЦА 1
Влияние FE и 1ADP на массу тела, потребление корма и коэффициент эффективности корма у крыс, получавших рационы, содержащие PN⋅HCl в дозе 7 мг / кг рациона или 0,7 мг / кг рациона в течение 5 недель 1